

Evidence for cadherin-11 cleavage in the synovium and partial characterization of its mechanism
- PMID: 25975695
- PMCID: PMC4449585
- DOI: 10.1186/s13075-015-0647-9
Evidence for cadherin-11 cleavage in the synovium and partial characterization of its mechanism
Abstract
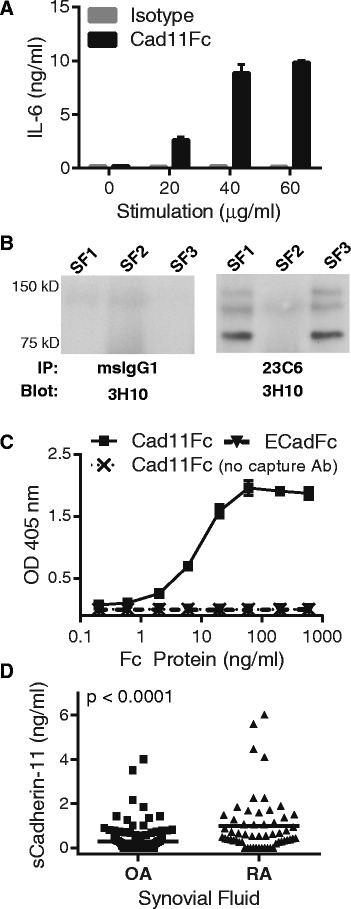
Introduction: Engagement of the homotypic cell-to-cell adhesion molecule cadherin-11 on rheumatoid arthritis (RA) synovial fibroblasts with a chimeric molecule containing the cadherin-11 extracellular binding domain stimulated cytokine, chemokine, and matrix metalloproteinases (MMP) release, implicating cadherin-11 signaling in RA pathogenesis. The objective of this study was to determine if cadherin-11 extracellular domain fragments are found inside the joint and if a physiologic synovial fibroblast cleavage pathway releases those fragments.
Methods: Cadherin-11 cleavage fragments were detected by western blot in cell media or lysates. Cleavage was interrupted using chemical inhibitors or short-interfering RNA (siRNA) gene silencing. The amount of cadherin-11 fragments in synovial fluid was measured by western blot and ELISA.
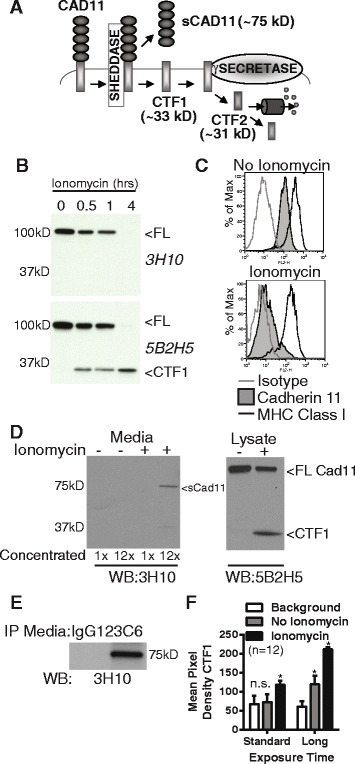
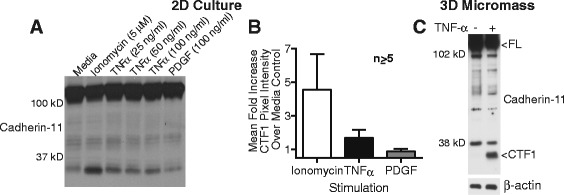
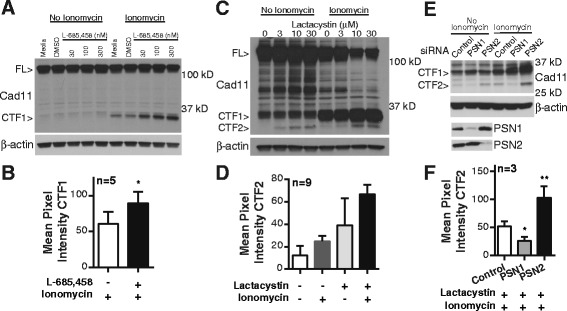
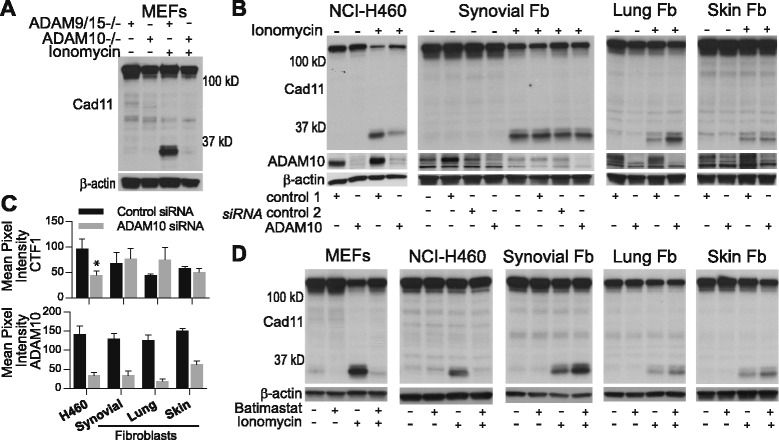
Results: Soluble cadherin-11 extracellular fragments were detected in human synovial fluid at significantly higher levels in RA samples compared to osteoarthritis (OA) samples. A cadherin-11 N-terminal extracellular binding domain fragment was shed from synovial fibroblasts after ionomycin stimulation, followed by presenilin 1 (PSN1)-dependent regulated intramembrane proteolysis of the retained membrane-bound C-terminal fragments. In addition to ionomycin-induced calcium flux, tumor necrosis factor (TNF)-α also stimulated cleavage in both two- and three-dimensional fibroblast cultures. Although cadherin-11 extracellular domains were shed by a disintegrin and metalloproteinase (ADAM) 10 in several cell types, a novel ADAM- and metalloproteinase-independent activity mediated shedding in primary human fibroblasts.
Conclusions: Cadherin-11 undergoes ectodomain shedding followed by regulated intramembrane proteolysis in synovial fibroblasts, triggered by a novel sheddase that generates extracelluar cadherin-11 fragments. Cadherin-11 fragments were enriched in RA synovial fluid, suggesting they may be a marker of synovial burden and may function to modify cadherin-11 interactions between synovial fibroblasts.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
