

An animal model of female adolescent cannabinoid exposure elicits a long-lasting deficit in presynaptic long-term plasticity
- PMID: 25979486
- PMCID: PMC4644105
- DOI: 10.1016/j.neuropharm.2015.04.034
An animal model of female adolescent cannabinoid exposure elicits a long-lasting deficit in presynaptic long-term plasticity
Abstract
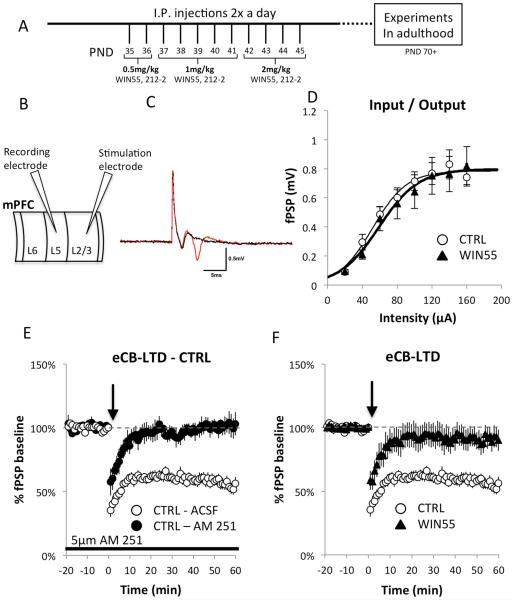
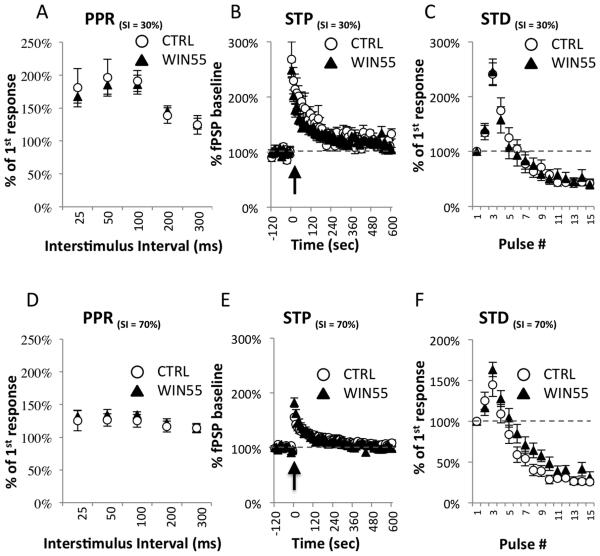
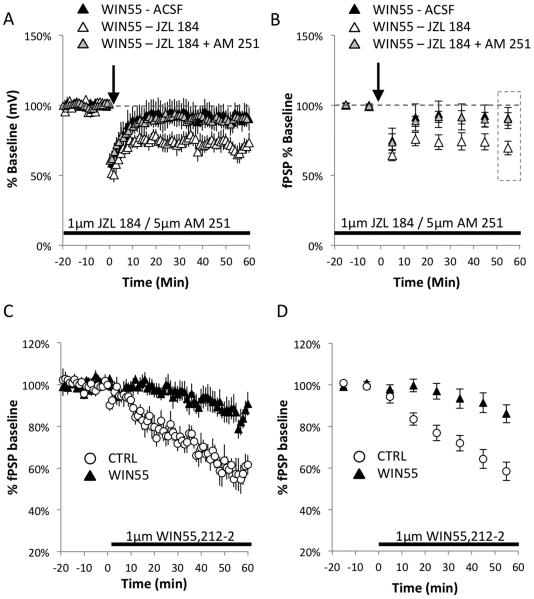
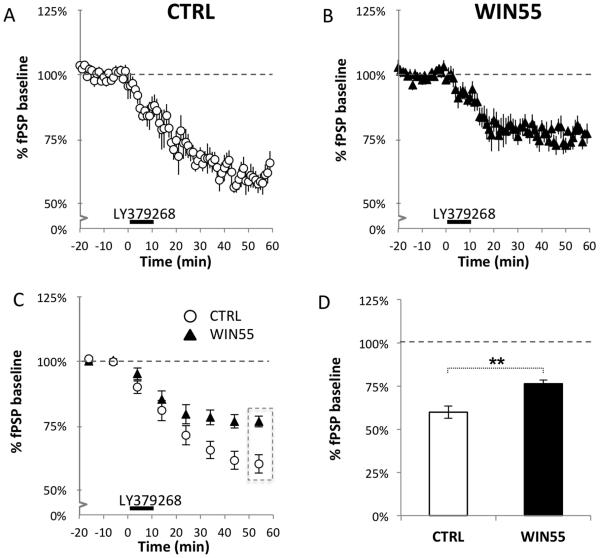
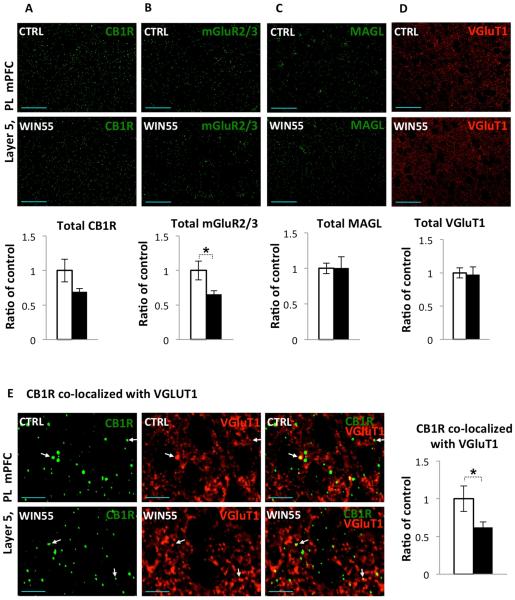
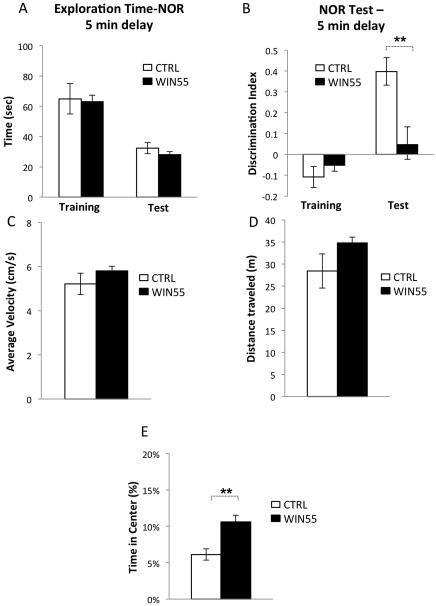
Cannabis continues to be the most accessible and popular illicit recreational drug. Whereas current data link adolescence cannabinoid exposure to increased risk for dependence on other drugs, depression, anxiety disorders and psychosis, the mechanism(s) underlying these adverse effects remains controversial. Here we show in a mouse model of female adolescent cannabinoid exposure deficient endocannabinoid (eCB)-mediated signaling and presynaptic forms of long-term depression at adult central glutamatergic synapses in the prefrontal cortex. Increasing endocannabinoid levels by blockade of monoacylglycerol lipase, the primary enzyme responsible for degrading the endocannabinoid 2-arachidonoylglycerol (2-AG), with the specific inhibitor JZL 184 ameliorates eCB-LTD deficits. The observed deficit in cortical presynaptic signaling may represent a neural maladaptation underlying network instability and abnormal cognitive functioning. Our study suggests that adolescent cannabinoid exposure may permanently impair brain functions, including the brain's intrinsic ability to appropriately adapt to external influences.
Keywords: Adolescent cannabis abuse; CB1R; Cannabinoids; LTD; mGluR2/3.
Published by Elsevier Ltd.
Figures
References
-
- Anderson PM, Pinault D, O'Brien TJ, Jones NC. Chronic administration of antipsychotics attenuates ongoing and ketamine-induced increases in cortical gamma oscillations. Int J Neuropsychopharmacol. 2014:1–10. - PubMed
-
- Arseneault L, Cannon M, Witton J, Murray RM. Causal association between cannabis and psychosis: examination of the evidence. Br J Psychiatry. 2004;184:110–117. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
