

Structural Insights into the Dynamic Process of β2-Adrenergic Receptor Signaling
- PMID: 25981665
- PMCID: PMC4441853
- DOI: 10.1016/j.cell.2015.04.043
Structural Insights into the Dynamic Process of β2-Adrenergic Receptor Signaling
Erratum in
- Cell. 2015 Sep 10;162(6):1431
Abstract
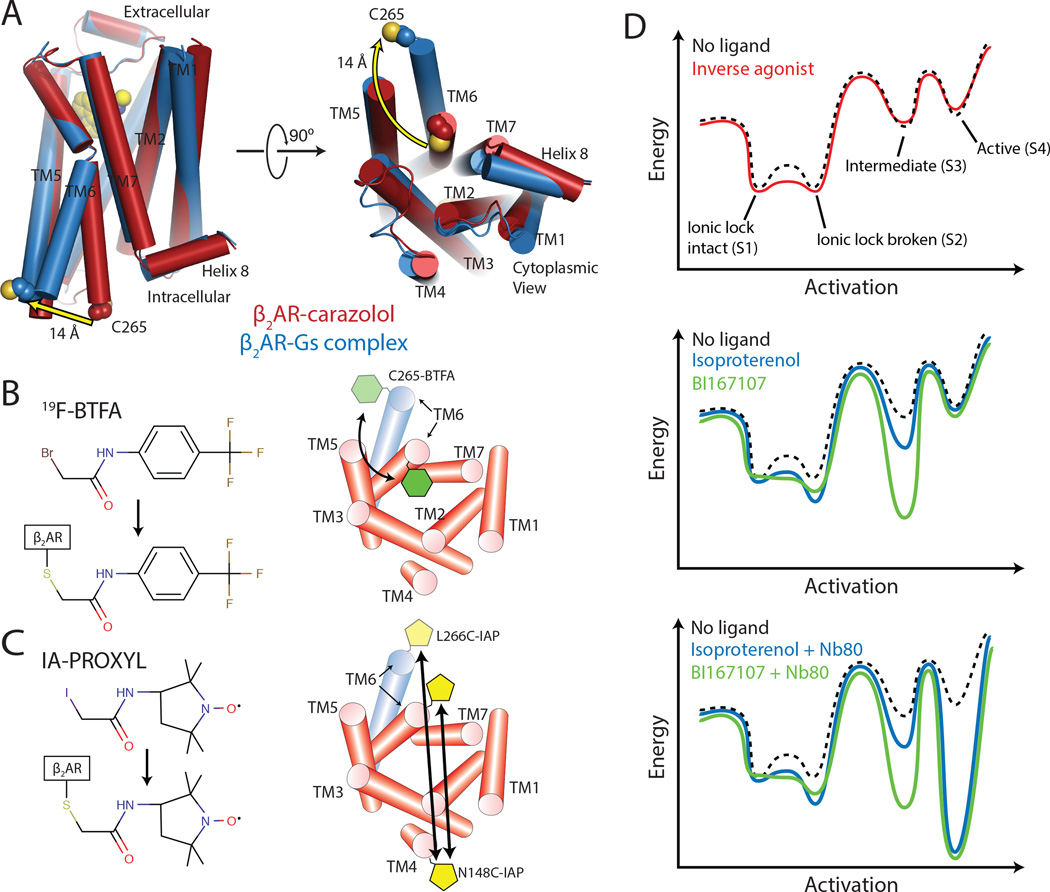
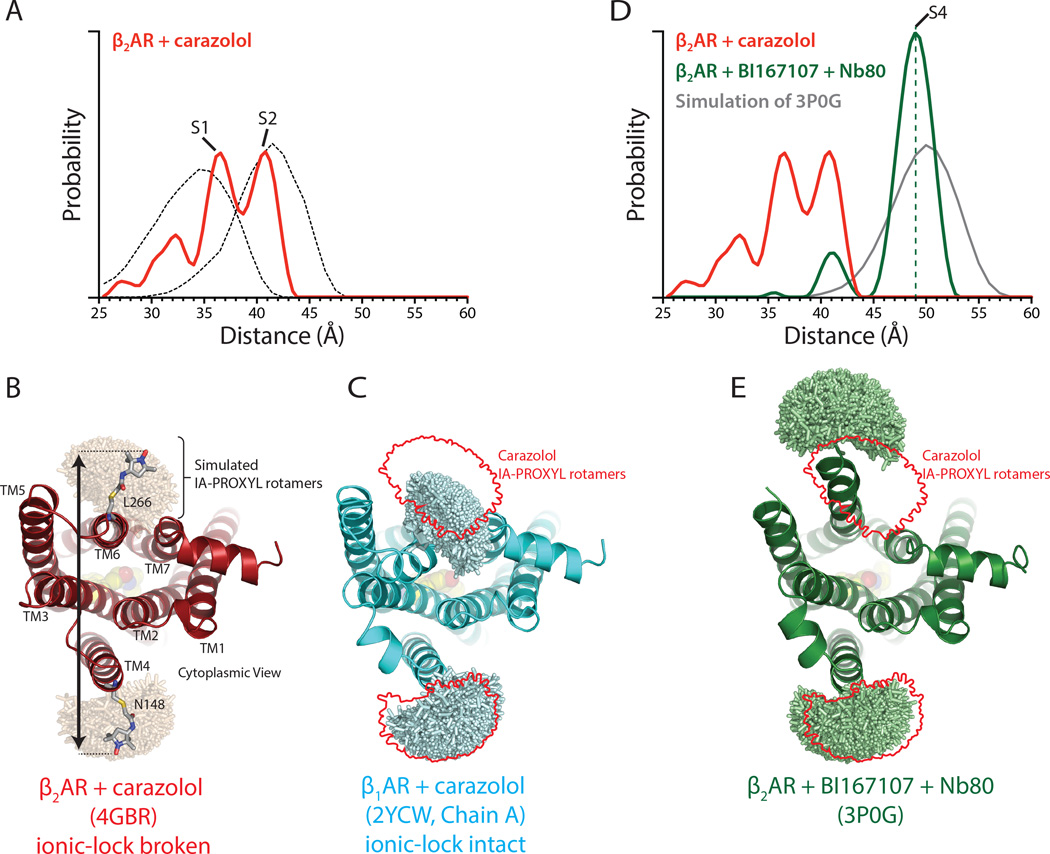
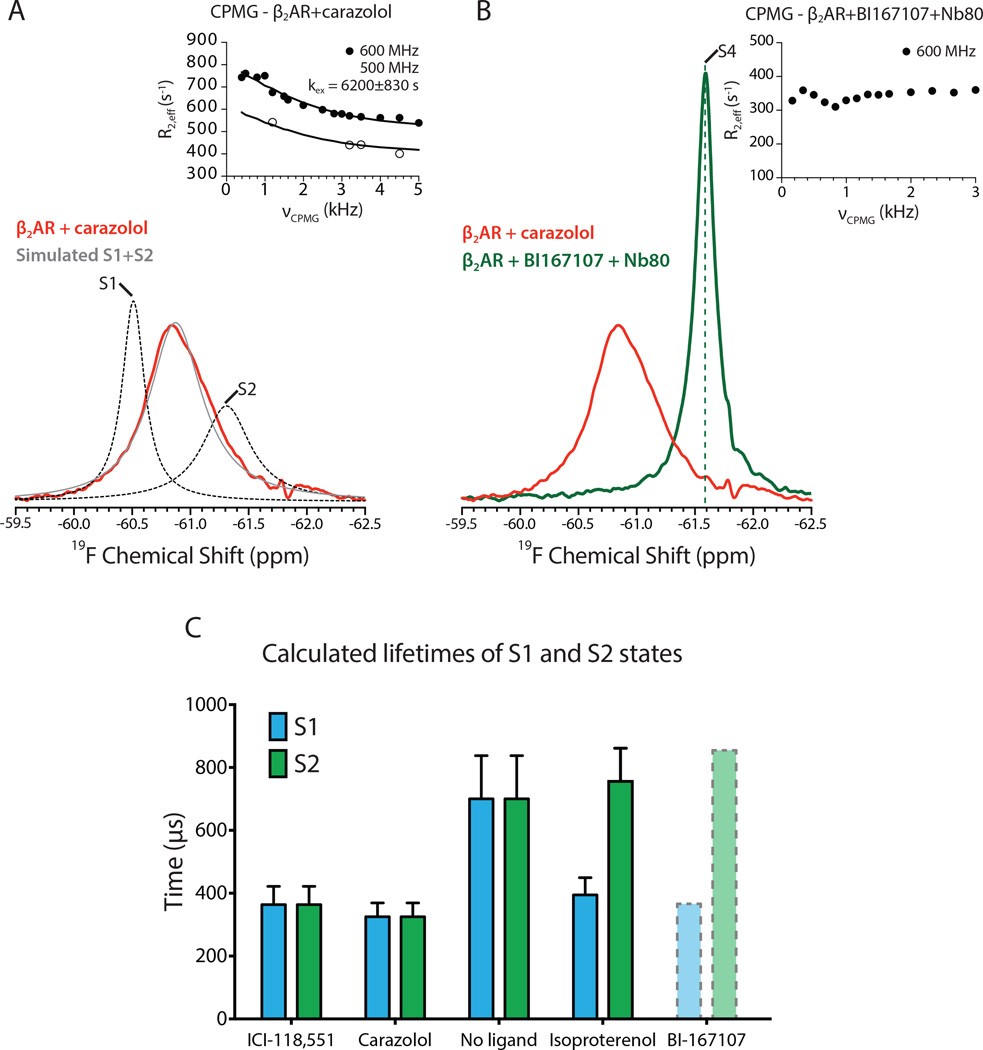
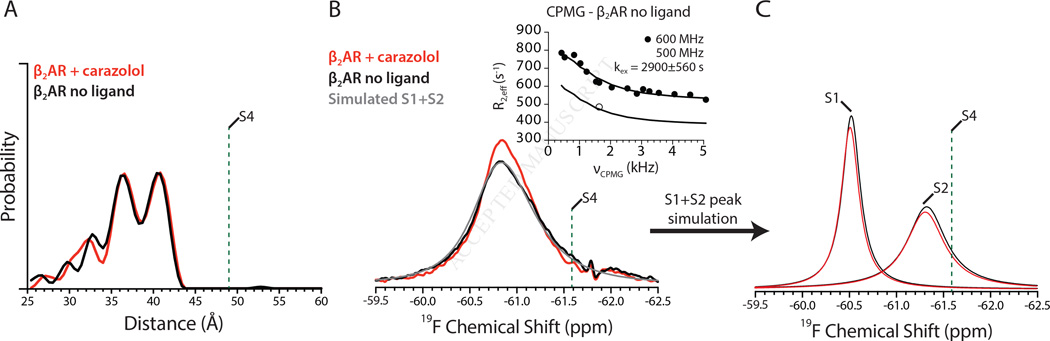
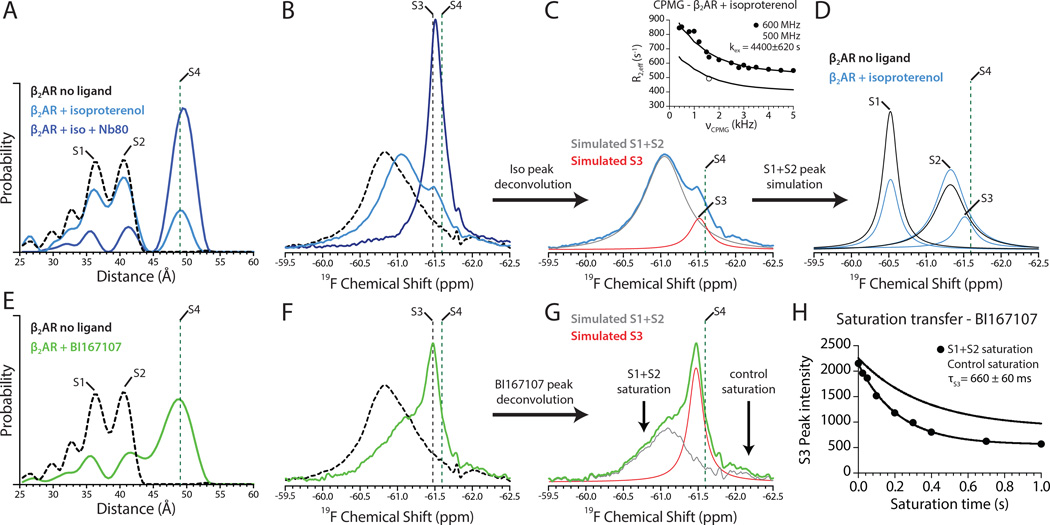
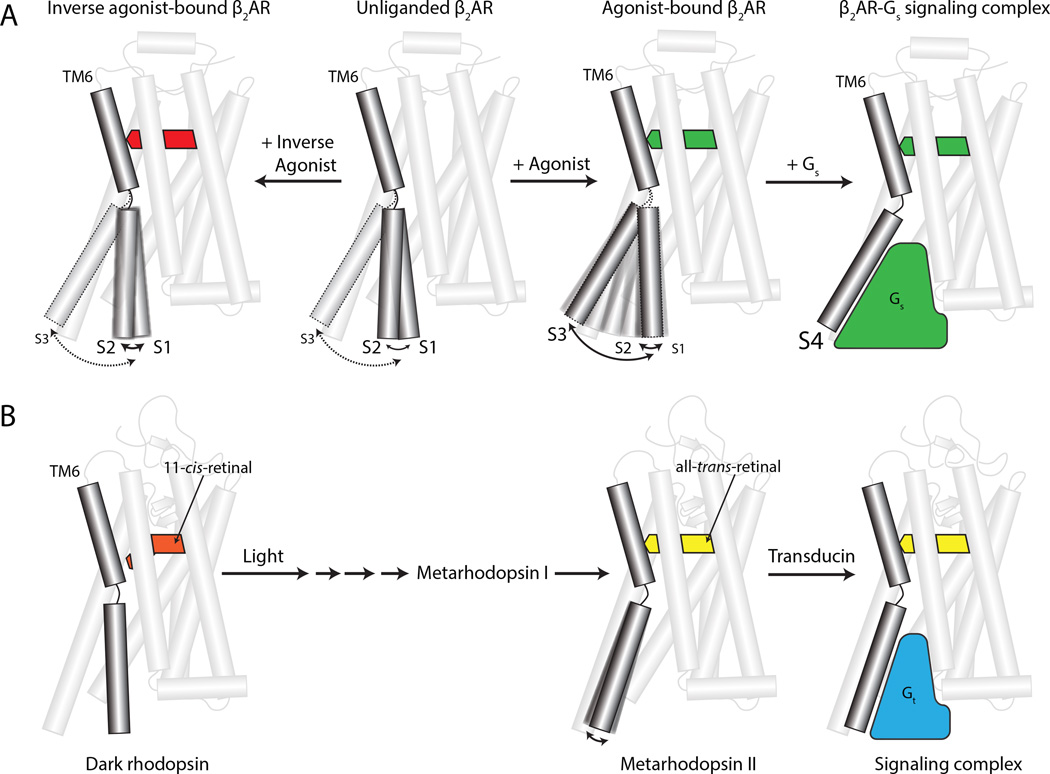
G-protein-coupled receptors (GPCRs) transduce signals from the extracellular environment to intracellular proteins. To gain structural insight into the regulation of receptor cytoplasmic conformations by extracellular ligands during signaling, we examine the structural dynamics of the cytoplasmic domain of the β2-adrenergic receptor (β2AR) using (19)F-fluorine NMR and double electron-electron resonance spectroscopy. These studies show that unliganded and inverse-agonist-bound β2AR exists predominantly in two inactive conformations that exchange within hundreds of microseconds. Although agonists shift the equilibrium toward a conformation capable of engaging cytoplasmic G proteins, they do so incompletely, resulting in increased conformational heterogeneity and the coexistence of inactive, intermediate, and active states. Complete transition to the active conformation requires subsequent interaction with a G protein or an intracellular G protein mimetic. These studies demonstrate a loose allosteric coupling of the agonist-binding site and G-protein-coupling interface that may generally be responsible for the complex signaling behavior observed for many GPCRs.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Baldwin AJ, Kay LE. NMR spectroscopy brings invisible protein states into focus. Nature chemical biology. 2009;5:808–814. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
