

Engineering strategies for the fermentative production of plant alkaloids in yeast
- PMID: 25981946
- PMCID: PMC4519383
- DOI: 10.1016/j.ymben.2015.05.001
Engineering strategies for the fermentative production of plant alkaloids in yeast
Abstract
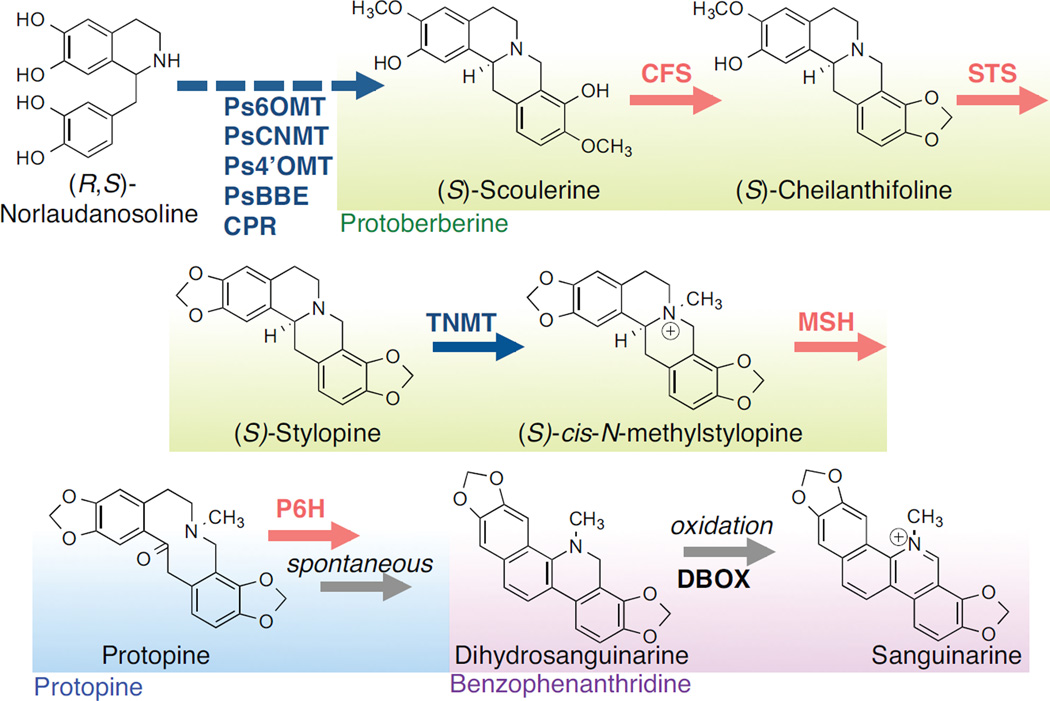
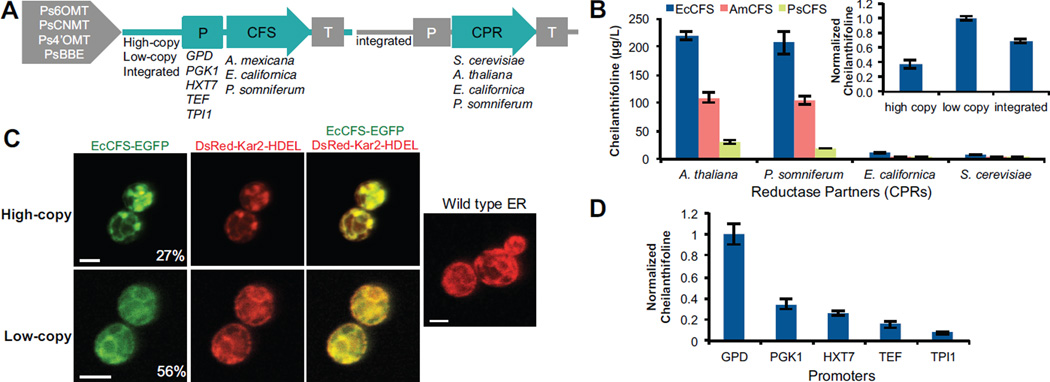
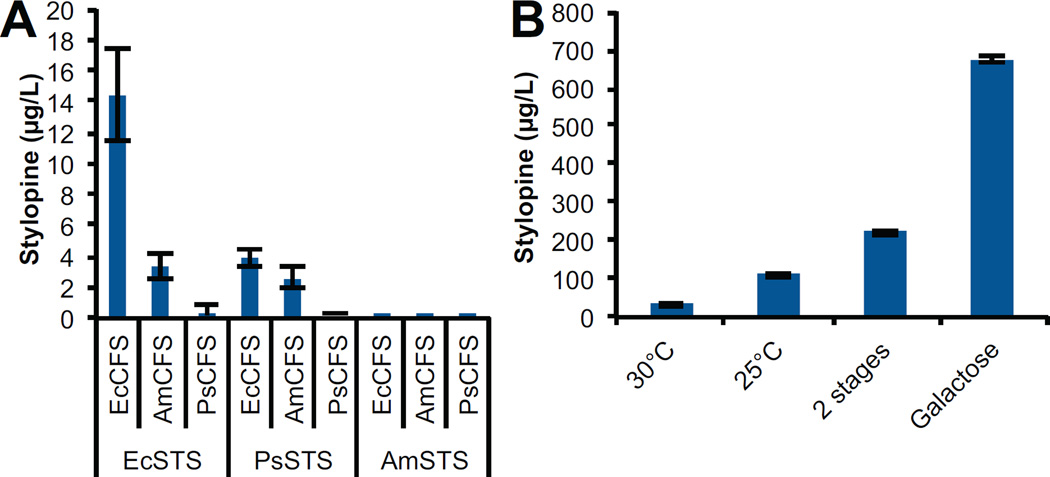
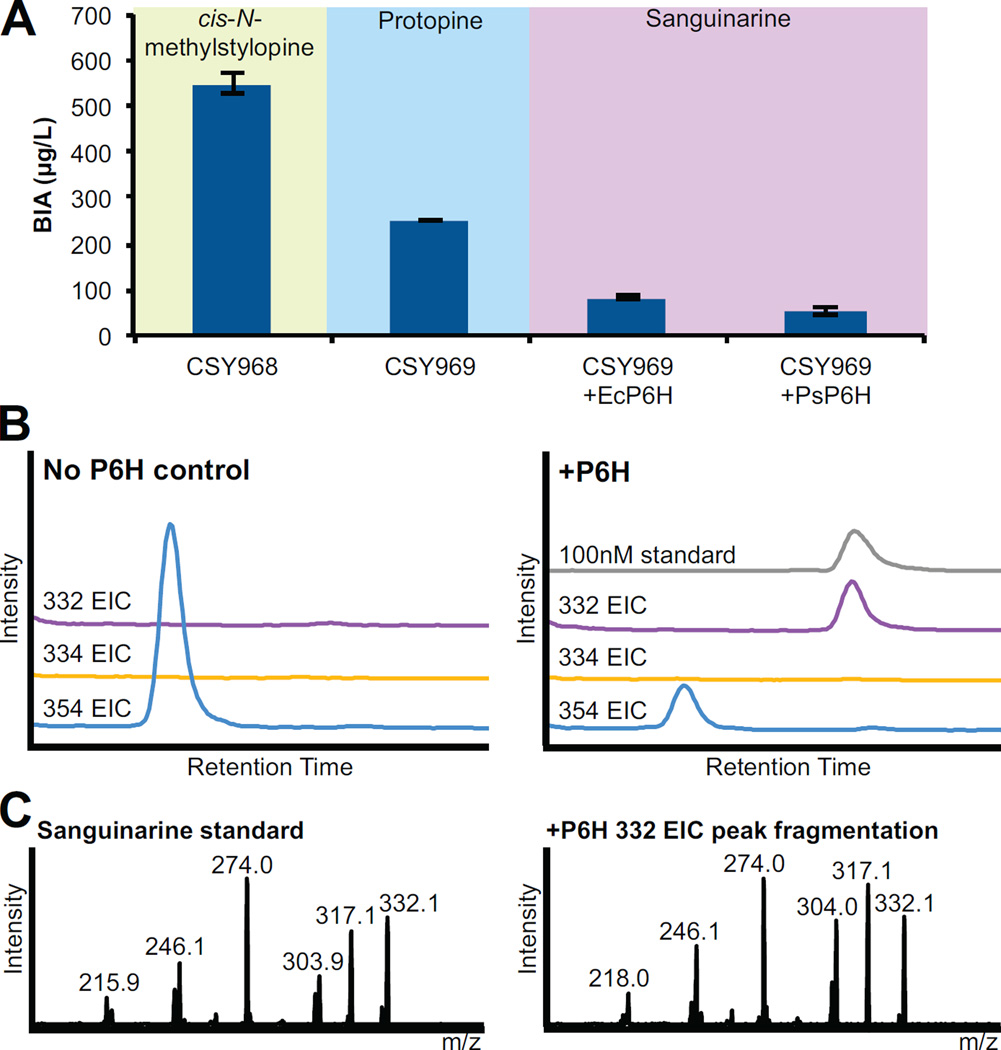
Microbial hosts engineered for the biosynthesis of plant natural products offer enormous potential as powerful discovery and production platforms. However, the reconstruction of these complex biosynthetic schemes faces numerous challenges due to the number of enzymatic steps and challenging enzyme classes associated with these pathways, which can lead to issues in metabolic load, pathway specificity, and maintaining flux to desired products. Cytochrome P450 enzymes are prevalent in plant specialized metabolism and are particularly difficult to express heterologously. Here, we describe the reconstruction of the sanguinarine branch of the benzylisoquinoline alkaloid pathway in Saccharomyces cerevisiae, resulting in microbial biosynthesis of protoberberine, protopine, and benzophenanthridine alkaloids through to the end-product sanguinarine, which we demonstrate can be efficiently produced in yeast in the absence of the associated biosynthetic enzyme. We achieved titers of 676 μg/L stylopine, 548 μg/L cis-N-methylstylopine, 252 μg/L protopine, and 80 μg/L sanguinarine from the engineered yeast strains. Through our optimization efforts, we describe genetic and culture strategies supporting the functional expression of multiple plant cytochrome P450 enzymes in the context of a large multi-step pathway. Our results also provided insight into relationships between cytochrome P450 activity and yeast ER physiology. We were able to improve the production of critical intermediates by 32-fold through genetic techniques and an additional 45-fold through culture optimization.
Keywords: Cytochrome P450; Plant natural products; Synthetic biology; Yeast.
Copyright © 2015 International Metabolic Engineering Society. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
Similar articles
-
Optimization of yeast-based production of medicinal protoberberine alkaloids.Microb Cell Fact. 2015 Sep 16;14:144. doi: 10.1186/s12934-015-0332-3. Microb Cell Fact. 2015. PMID: 26376732 Free PMC article.
-
Complete biosynthesis of the bisbenzylisoquinoline alkaloids guattegaumerine and berbamunine in yeast.Proc Natl Acad Sci U S A. 2021 Dec 21;118(51):e2112520118. doi: 10.1073/pnas.2112520118. Proc Natl Acad Sci U S A. 2021. PMID: 34903659 Free PMC article.
-
Plug-and-Play Benzylisoquinoline Alkaloid Biosynthetic Gene Discovery in Engineered Yeast.Methods Enzymol. 2016;575:143-78. doi: 10.1016/bs.mie.2016.03.023. Epub 2016 Apr 25. Methods Enzymol. 2016. PMID: 27417928
-
Engineering Critical Enzymes and Pathways for Improved Triterpenoid Biosynthesis in Yeast.ACS Synth Biol. 2020 Sep 18;9(9):2214-2227. doi: 10.1021/acssynbio.0c00124. Epub 2020 Aug 24. ACS Synth Biol. 2020. PMID: 32786348 Review.
-
Yeast factories for the production of aromatic compounds: from building blocks to plant secondary metabolites.J Ind Microbiol Biotechnol. 2016 Nov;43(11):1611-1624. doi: 10.1007/s10295-016-1824-9. Epub 2016 Aug 31. J Ind Microbiol Biotechnol. 2016. PMID: 27581441 Review.
Cited by
-
Engineering biosynthesis of the anticancer alkaloid noscapine in yeast.Nat Commun. 2016 Jul 5;7:12137. doi: 10.1038/ncomms12137. Nat Commun. 2016. PMID: 27378283 Free PMC article.
-
Complete biosynthesis of opioids in yeast.Science. 2015 Sep 4;349(6252):1095-100. doi: 10.1126/science.aac9373. Epub 2015 Aug 13. Science. 2015. PMID: 26272907 Free PMC article.
-
Optimization of yeast-based production of medicinal protoberberine alkaloids.Microb Cell Fact. 2015 Sep 16;14:144. doi: 10.1186/s12934-015-0332-3. Microb Cell Fact. 2015. PMID: 26376732 Free PMC article.
-
Engineering Electron Transfer Pathway of Cytochrome P450s.Molecules. 2024 May 24;29(11):2480. doi: 10.3390/molecules29112480. Molecules. 2024. PMID: 38893355 Free PMC article. Review.
-
De novo synthesis of 1-phenethylisoquinoline in engineered Escherichia coli.Synth Syst Biotechnol. 2024 Nov 9;10(1):271-280. doi: 10.1016/j.synbio.2024.10.007. eCollection 2025. Synth Syst Biotechnol. 2024. PMID: 39650803 Free PMC article.
References
-
- Bauer W, Zenk M. Two methylenedioxy bridge forming cytochrome P-450 dependent enzymes are involved in (S)-stylopine biosynthesis. Phytochemistry. 1991:30.
-
- Beaudoin GW, Facchini PJ. Isolation and characterization of a cDNA encoding (S)-cis-N-methylstylopine 14-hydroxylase from opium poppy, a key enzyme in sanguinarine biosynthesis. Biochem. Biophys. Res. Commun. 2013;431:597–603. - PubMed
-
- Bevis BJ, Hammond AT, Reinke CA, Glick BS. De novo formation of transitional ER sites and Golgi structures in Pichia pastoris. Nat. Cell Biol. 2002;4:750–756. - PubMed
-
- Chemler JA, Koffas MG. Metabolic engineering for plant natural product biosynthesis in microbes. Curr. Opin. Biotechnol. 2008;19:597–605. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
