

The mechanism of inhibition of protein synthesis by the proline-rich peptide oncocin
- PMID: 25984972
- PMCID: PMC4456192
- DOI: 10.1038/nsmb.3031
The mechanism of inhibition of protein synthesis by the proline-rich peptide oncocin
Abstract
Antibiotic-resistant bacteria are a global health issue necessitating the development of new effective therapeutics. Proline-rich antimicrobial peptides (PrAMPs), which include oncocins, are an extensively studied class of AMPs that counteract bacterial infection at submicromolar concentrations. Oncocins enter and kill bacteria by inhibiting certain targets rather than by acting through membrane lysis. Although they have recently been reported to bind DnaK and the bacterial ribosome, their mode of inhibition has remained elusive. Here we report the crystal structure of the oncocin derivative Onc112 bound to the Thermus thermophilus 70S ribosome. Strikingly, this 19-residue proline-rich peptide manifests the features of several known classes of ribosome inhibitors by simultaneously blocking the peptidyl transferase center and the peptide-exit tunnel of the ribosome. This high-resolution structure thus reveals the mechanism by which oncocins inhibit protein synthesis, providing an opportunity for structure-based design of new-generation therapeutics.
Figures
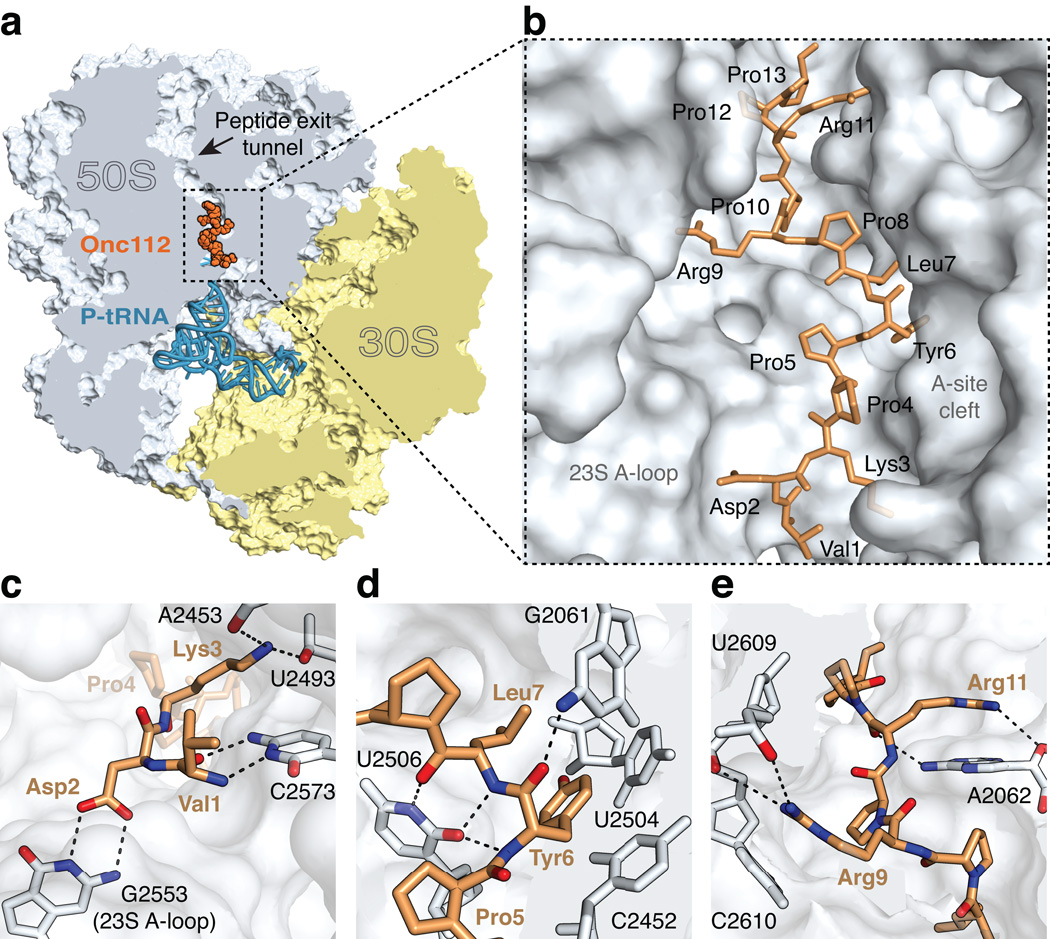
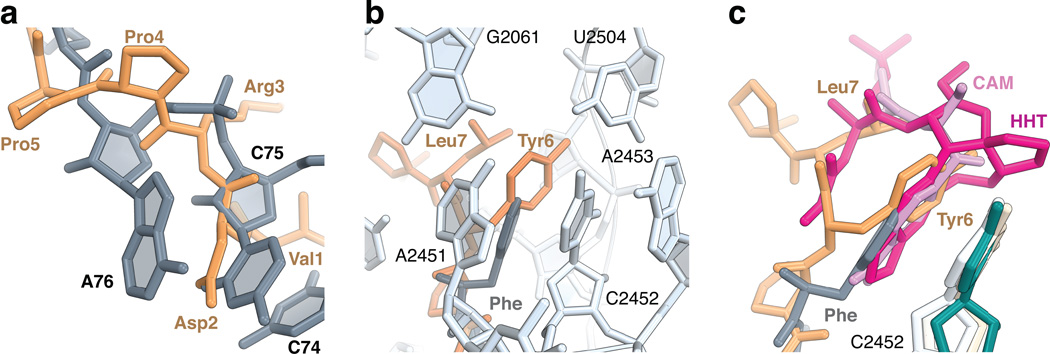
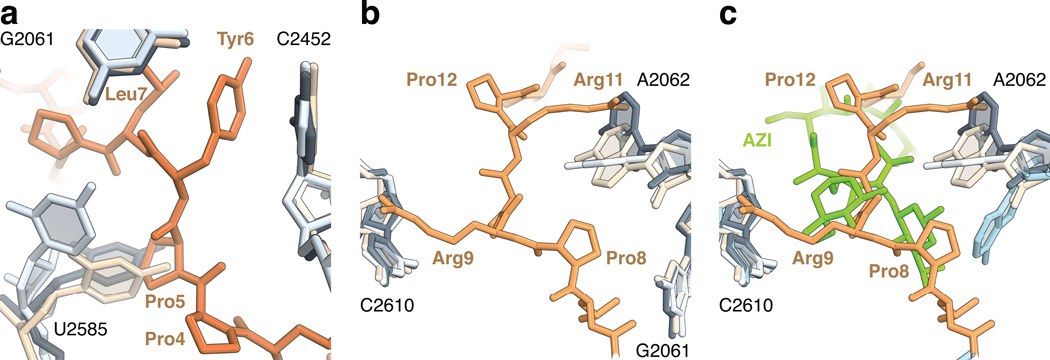
References
-
- de Souza Candido E, et al. The use of versatile plant antimicrobial peptides in agribusiness and human health. Peptides. 2014;55:65–78. - PubMed
-
- Li W, et al. Proline-rich antimicrobial peptides: potential therapeutics against antibiotic-resistant bacteria. Amino Acids. 2014;46:2287–2294. - PubMed
-
- Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415:389–395. - PubMed
-
- Brogden KA, Ackermann M, McCray PB, Jr, Tack BF. Antimicrobial peptides in animals and their role in host defences. Int J Antimicrob Agents. 2003;22:465–478. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
