

Genome-wide maps of recombination and chromosome segregation in human oocytes and embryos show selection for maternal recombination rates
- PMID: 25985139
- PMCID: PMC4770575
- DOI: 10.1038/ng.3306
Genome-wide maps of recombination and chromosome segregation in human oocytes and embryos show selection for maternal recombination rates
Abstract
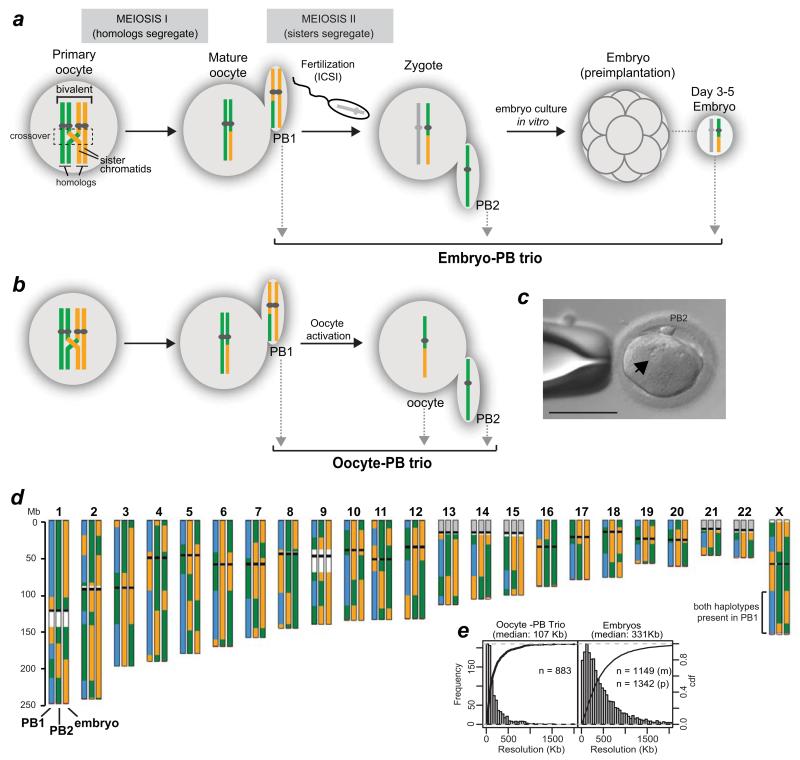
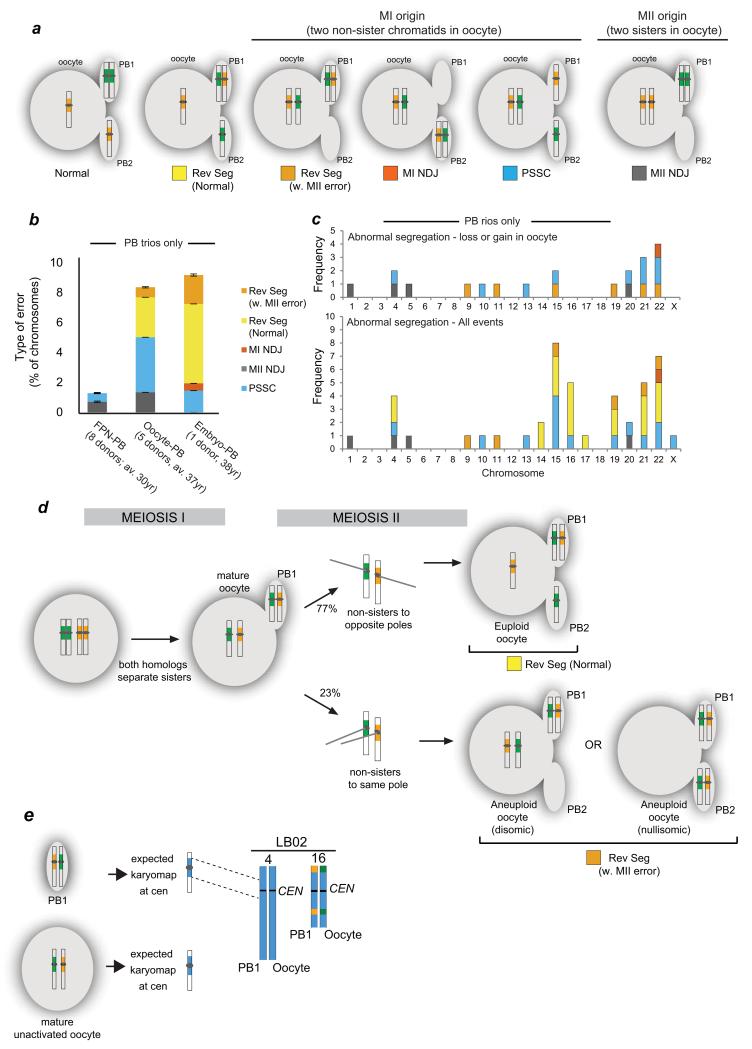
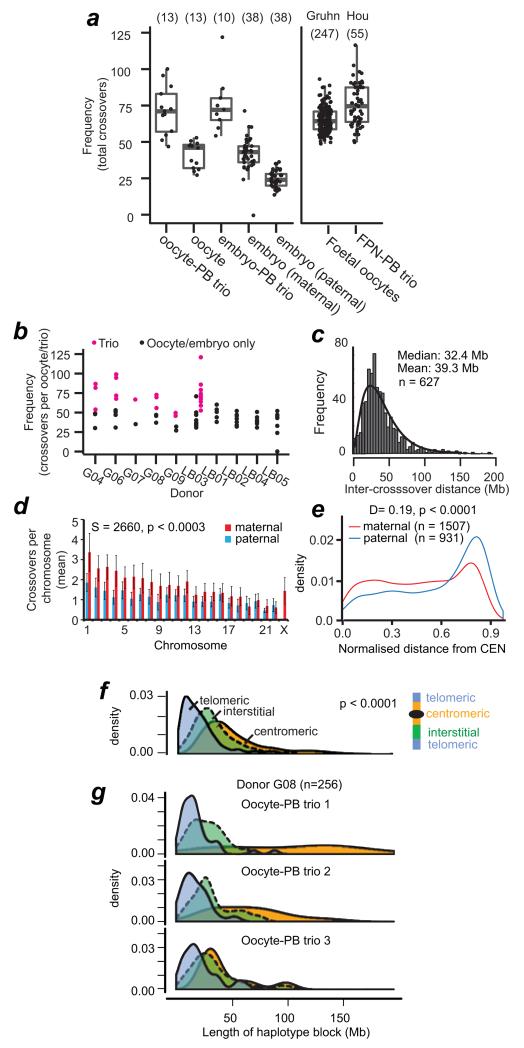
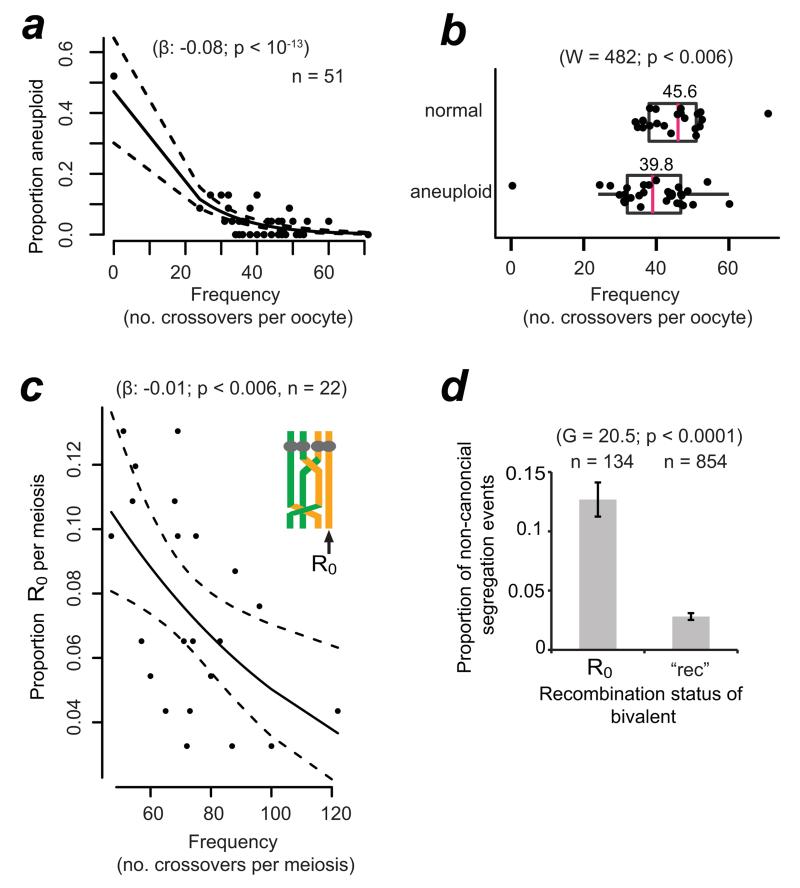
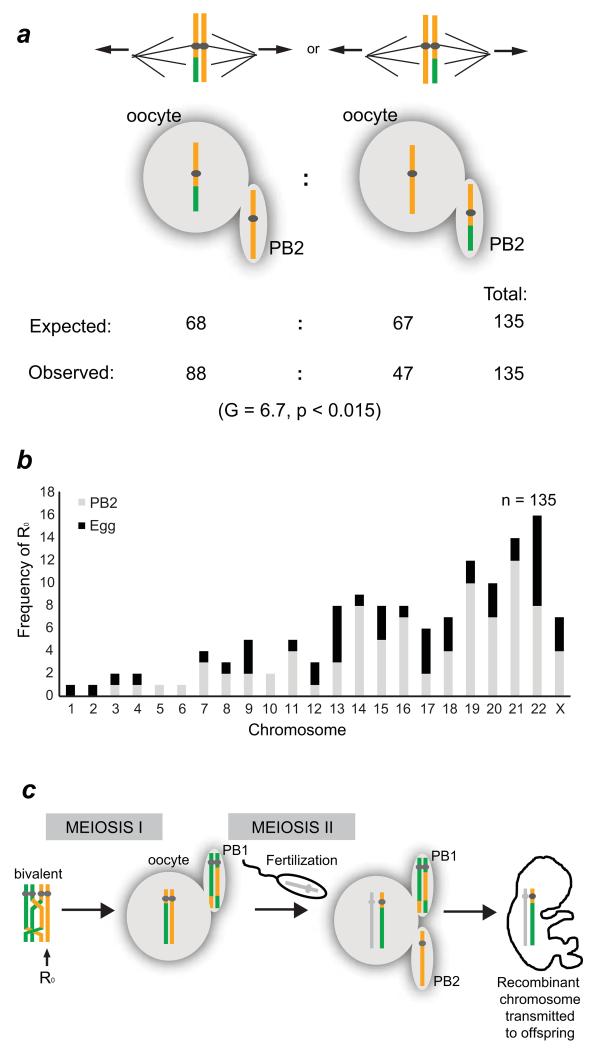
Crossover recombination reshuffles genes and prevents errors in segregation that lead to extra or missing chromosomes (aneuploidy) in human eggs, a major cause of pregnancy failure and congenital disorders. Here we generate genome-wide maps of crossovers and chromosome segregation patterns by recovering all three products of single female meioses. Genotyping >4 million informative SNPs from 23 complete meioses allowed us to map 2,032 maternal and 1,342 paternal crossovers and to infer the segregation patterns of 529 chromosome pairs. We uncover a new reverse chromosome segregation pattern in which both homologs separate their sister chromatids at meiosis I; detect selection for higher recombination rates in the female germ line by the elimination of aneuploid embryos; and report chromosomal drive against non-recombinant chromatids at meiosis II. Collectively, our findings show that recombination not only affects homolog segregation at meiosis I but also the fate of sister chromatids at meiosis II.
Figures
Comment in
-
Double trouble in human aneuploidy.Nat Genet. 2015 Jul;47(7):696-8. doi: 10.1038/ng.3344. Nat Genet. 2015. PMID: 26111508
-
Chromosome segregation: human female meiosis breaks all the rules.Curr Biol. 2015 Aug 3;25(15):R654-6. doi: 10.1016/j.cub.2015.06.054. Curr Biol. 2015. PMID: 26241139
References
-
- Hassold T, Hunt P. To err (meiotically) is human: the genesis of human aneuploidy. Nat Rev Genet. 2001;2:280–291. - PubMed
-
- Zaragoza MV, et al. Nondisjunction of human acrocentric chromosomes: studies of 432 trisomic fetuses and liveborns. Hum Genet. 1994;94:411–417. - PubMed
-
- Hassold TJ, Jacobs PA. Trisomy in man. Annu Rev Genet. 1984;18:69–97. - PubMed
-
- Hassold T, et al. A cytogenetic study of 1000 spontaneous abortions. Ann Hum Genet. 1980;44:151–178. - PubMed
Methods-only references
-
- Kuwayama M. Highly efficient vitrification for cryopreservation of human oocytes and embryos: the Cryotop method. Theriogenology. 2007;67:73–80. - PubMed
-
- Kuwayama M, Vajta G, Kato O, Leibo SP. Highly efficient vitrification method for cryopreservation of human oocytes. Reprod Biomed Online. 2005;11:300–308. - PubMed
-
- Wickham H. ggplot2: Elegant Graphics for Data Analysis. Use R. 2009:1–212.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
