

Genome-wide patterns and properties of de novo mutations in humans
- PMID: 25985141
- PMCID: PMC4485564
- DOI: 10.1038/ng.3292
Genome-wide patterns and properties of de novo mutations in humans
Abstract
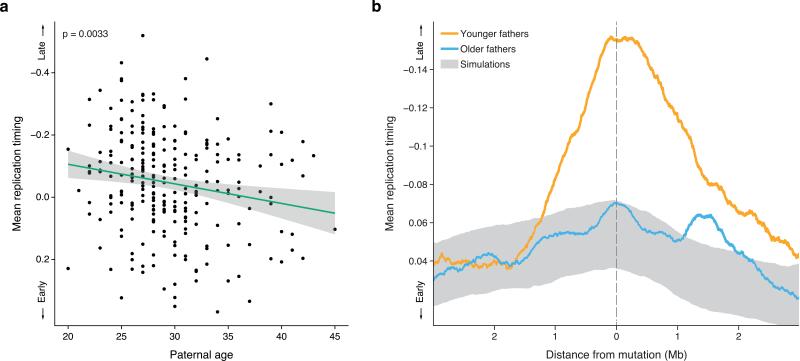
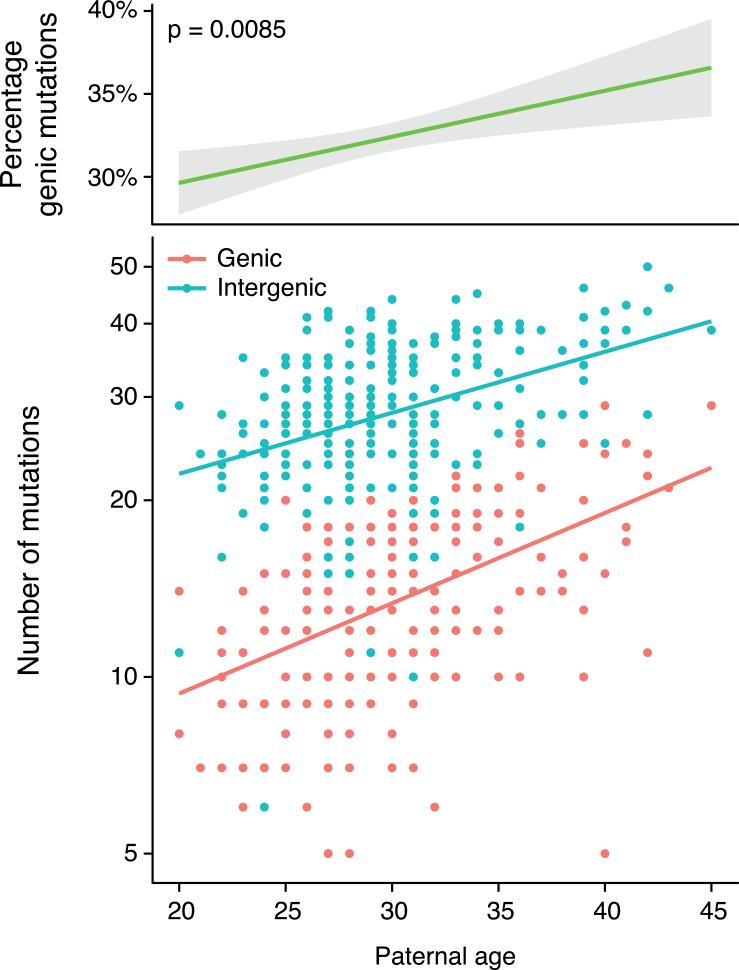
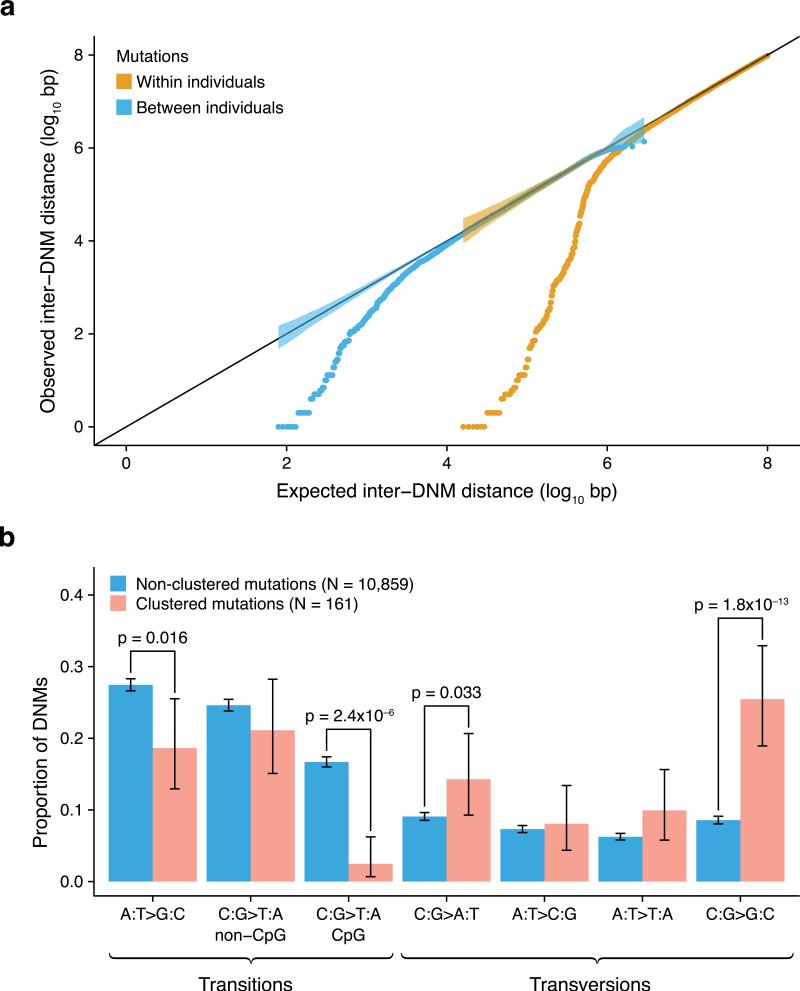
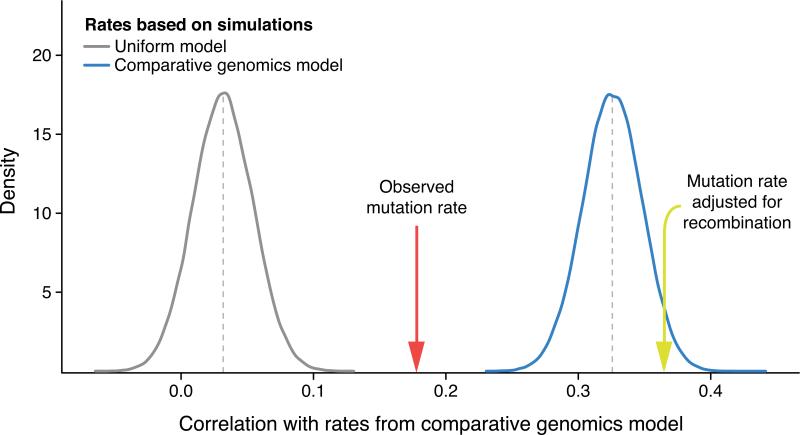
Mutations create variation in the population, fuel evolution and cause genetic diseases. Current knowledge about de novo mutations is incomplete and mostly indirect. Here we analyze 11,020 de novo mutations from the whole genomes of 250 families. We show that de novo mutations in the offspring of older fathers are not only more numerous but also occur more frequently in early-replicating, genic regions. Functional regions exhibit higher mutation rates due to CpG dinucleotides and show signatures of transcription-coupled repair, whereas mutation clusters with a unique signature point to a new mutational mechanism. Mutation and recombination rates independently associate with nucleotide diversity, and regional variation in human-chimpanzee divergence is only partly explained by heterogeneity in mutation rate. Finally, we provide a genome-wide mutation rate map for medical and population genetics applications. Our results provide new insights and refine long-standing hypotheses about human mutagenesis.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
