

Mechanisms of clonal evolution in childhood acute lymphoblastic leukemia
- PMID: 25985233
- PMCID: PMC4475638
- DOI: 10.1038/ni.3160
Mechanisms of clonal evolution in childhood acute lymphoblastic leukemia
Erratum in
-
Author Correction: Mechanisms of clonal evolution in childhood acute lymphoblastic leukemia.Nat Immunol. 2026 Feb;27(2):376. doi: 10.1038/s41590-025-02375-5. Nat Immunol. 2026. PMID: 41315083 No abstract available.
Abstract
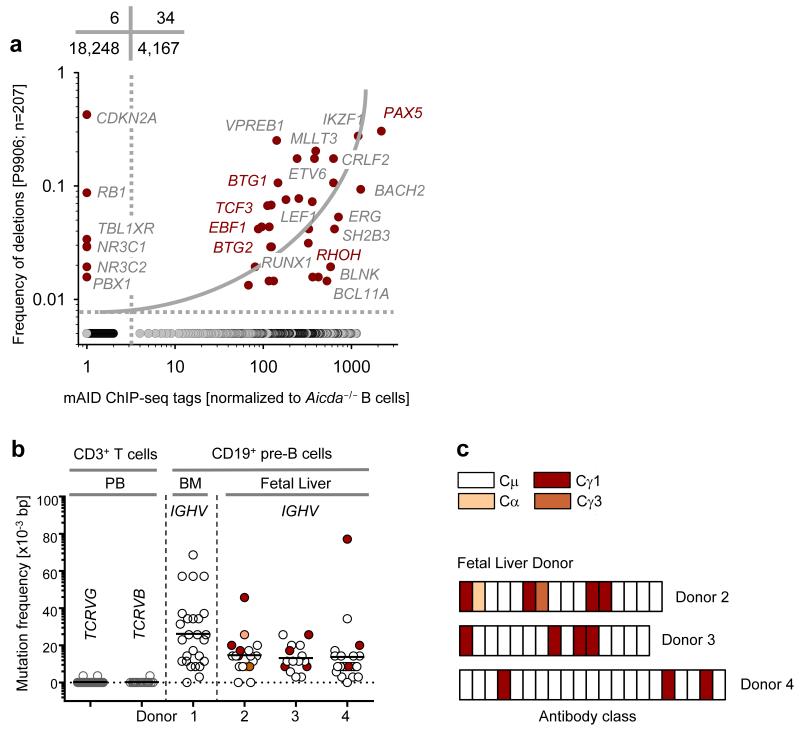
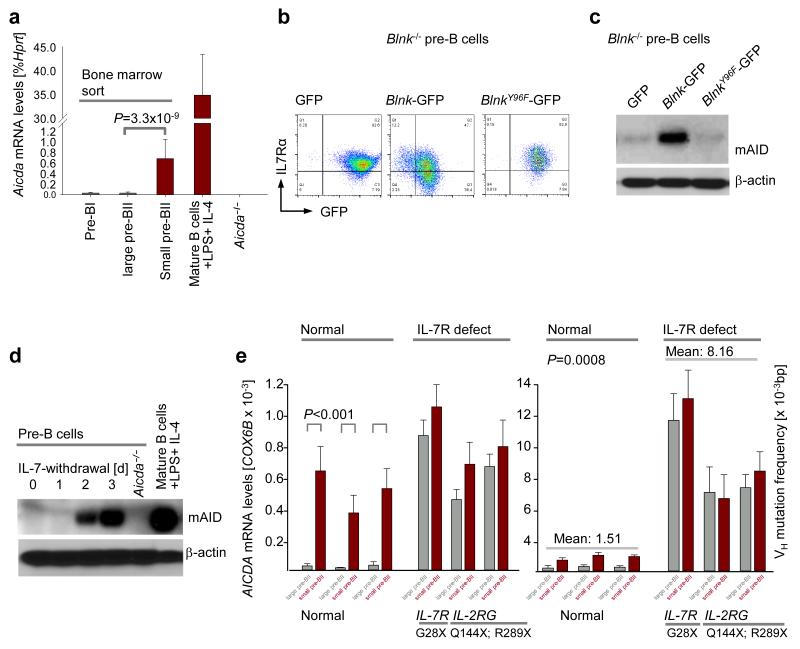
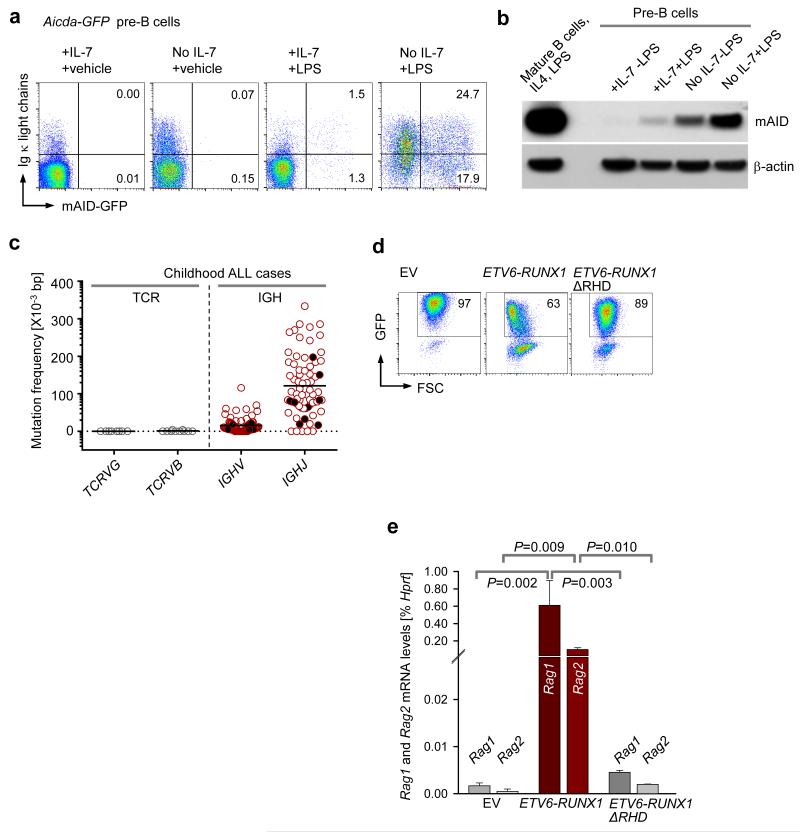
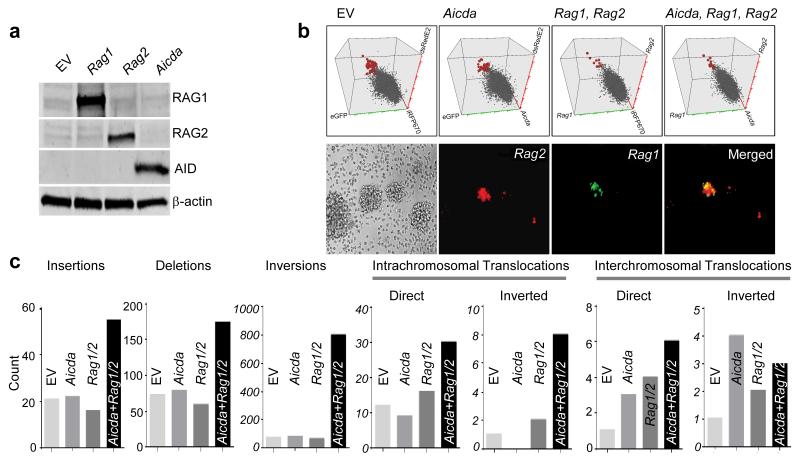
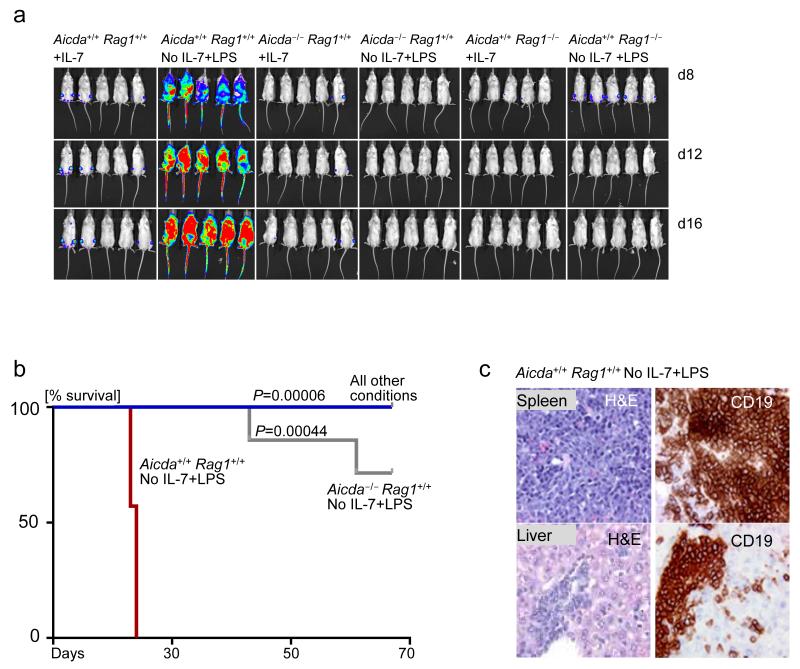
Childhood acute lymphoblastic leukemia (ALL) can often be traced to a pre-leukemic clone carrying a prenatal genetic lesion. Postnatally acquired mutations then drive clonal evolution toward overt leukemia. The enzymes RAG1-RAG2 and AID, which diversify immunoglobulin-encoding genes, are strictly segregated in developing cells during B lymphopoiesis and peripheral mature B cells, respectively. Here we identified small pre-BII cells as a natural subset with increased genetic vulnerability owing to concurrent activation of these enzymes. Consistent with epidemiological findings on childhood ALL etiology, susceptibility to genetic lesions during B lymphopoiesis at the transition from the large pre-BII cell stage to the small pre-BII cell stage was exacerbated by abnormal cytokine signaling and repetitive inflammatory stimuli. We demonstrated that AID and RAG1-RAG2 drove leukemic clonal evolution with repeated exposure to inflammatory stimuli, paralleling chronic infections in childhood.
Figures

References
-
- Wiemels JL, et al. Prenatal origin of acute lymphoblastic leukaemia in children. Lancet. 1999;354:1499–1503. - PubMed
-
- Greaves MF, Wiemels J. Origins of chromosome translocations in childhood leukaemia. Nat. Rev. Cancer. 2003;3:639–649. - PubMed
-
- Bateman CM, et al. Acquisition of genome-wide copy number alterations in monozygotic twins with acute lymphoblastic leukemia. Blood. 2010;115:3553–3558. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
