

Sphingosine 1-phosphate is a ligand for peroxisome proliferator-activated receptor-γ that regulates neoangiogenesis
- PMID: 25985799
- PMCID: PMC6137673
- DOI: 10.1096/fj.14-261289
Sphingosine 1-phosphate is a ligand for peroxisome proliferator-activated receptor-γ that regulates neoangiogenesis
Abstract
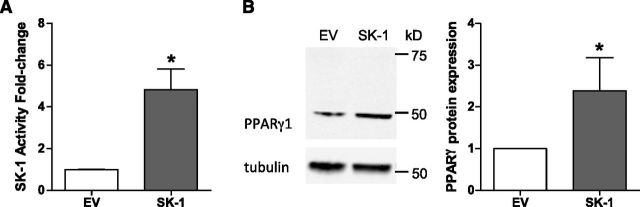
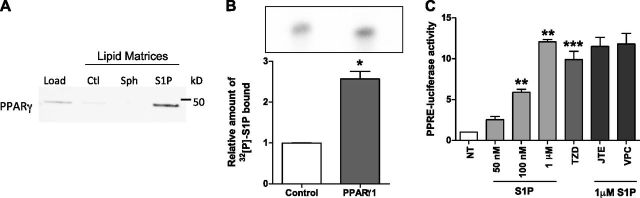
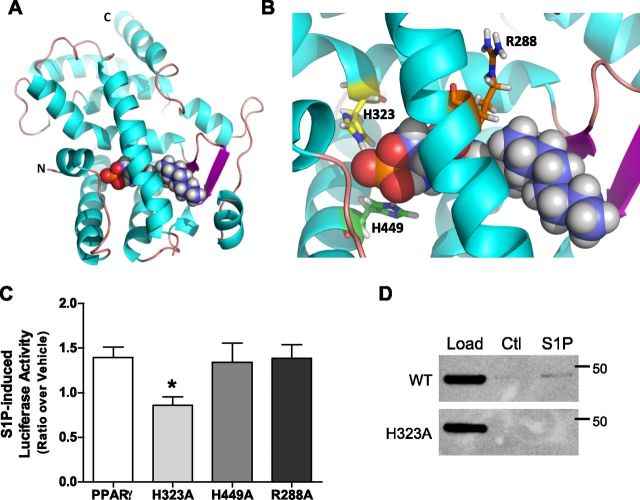
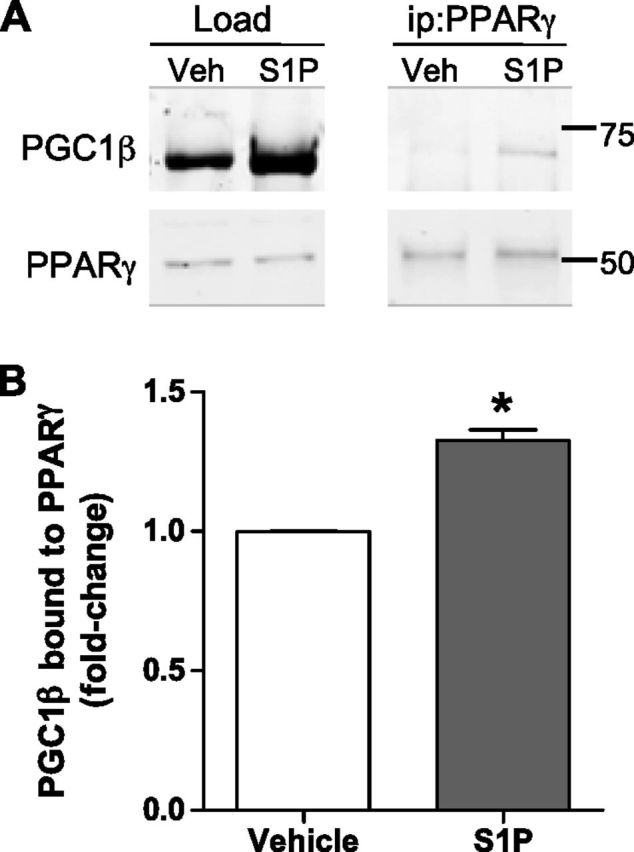
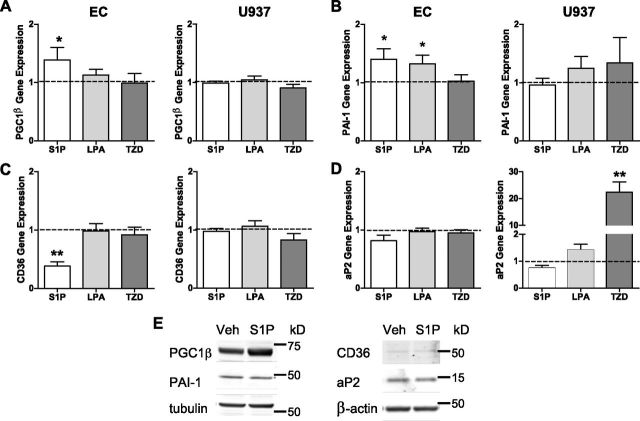
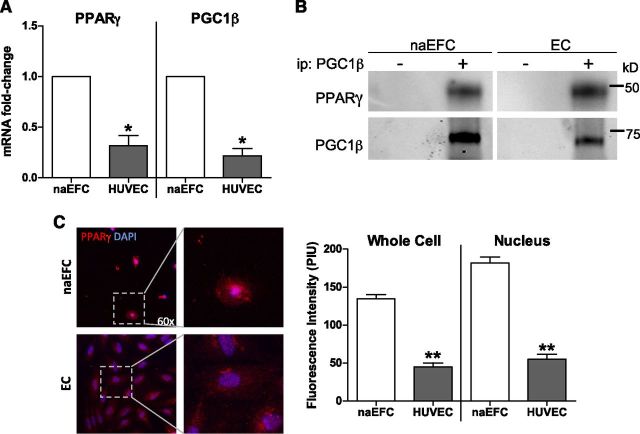
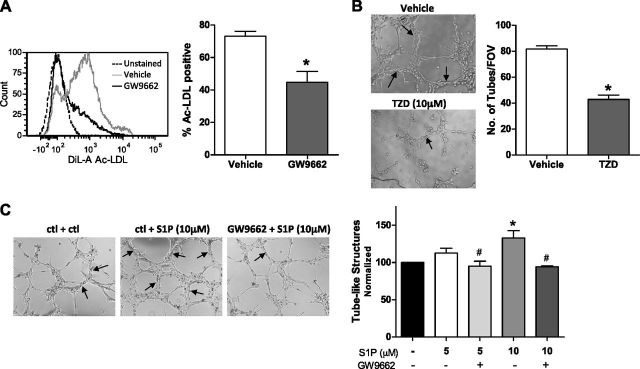
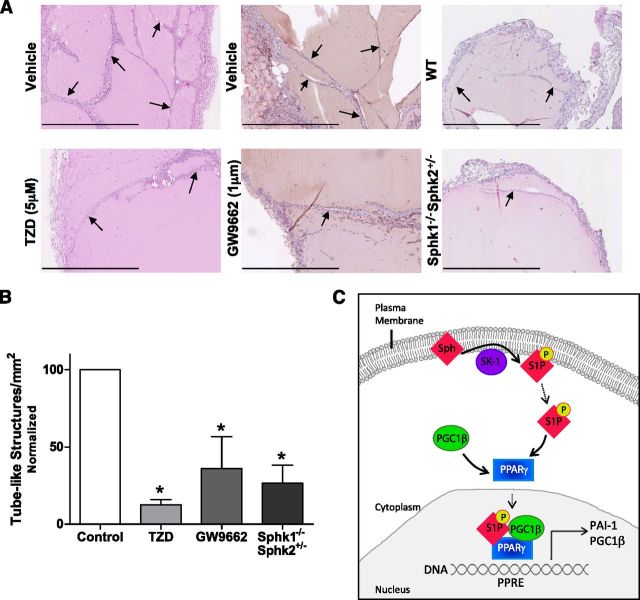
Sphingosine 1-phosphate (S1P) is a bioactive lipid that can function both extracellularly and intracellularly to mediate a variety of cellular processes. Using lipid affinity matrices and a radiolabeled lipid binding assay, we reveal that S1P directly interacts with the transcription factor peroxisome proliferator-activated receptor (PPAR)γ. Herein, we show that S1P treatment of human endothelial cells (ECs) activated a luciferase-tagged PPARγ-specific gene reporter by ∼12-fold, independent of the S1P receptors. More specifically, in silico docking, gene reporter, and binding assays revealed that His323 of the PPARγ ligand binding domain is important for binding to S1P. PPARγ functions when associated with coregulatory proteins, and herein we identify that peroxisome proliferator-activated receptor-γ coactivator 1 (PGC1)β binds to PPARγ in ECs and their progenitors (nonadherent endothelial forming cells) and that the formation of this PPARγ:PGC1β complex is increased in response to S1P. ECs treated with S1P selectively regulated known PPARγ target genes with PGC1β and plasminogen-activated inhibitor-1 being increased, no change to adipocyte fatty acid binding protein 2 and suppression of CD36. S1P-induced in vitro tube formation was significantly attenuated in the presence of the PPARγ antagonist GW9662, and in vivo application of GW9662 also reduced vascular development in Matrigel plugs. Interestingly, activation of PPARγ by the synthetic ligand troglitazone also reduced tube formation in vitro and in vivo. To support this, Sphk1(-/-)Sphk2(+/-) mice, with low circulating S1P levels, demonstrated a similar reduction in vascular development. Taken together, our data reveal that the transcription factor, PPARγ, is a bona fide intracellular target for S1P and thus suggest that the S1P:PPARγ:PGC1β complex may be a useful target to manipulate neovascularization.
Keywords: endothelial; neovascularization; transcription factor.
© FASEB.
Conflict of interest statement
The authors thank Dr. David Dimasi for preparing the ECs, Samantha Escarbe for preparing the nonadherent endothelial forming cells, Lorena Davies for technical assistance with sphingosine 1-phosphate analysis, Dr. James Fells for his assistance with the
Figures
References
-
- Pitson S. M. (2011) Regulation of sphingosine kinase and sphingolipid signaling. Trends Biochem. Sci. , 97–107 - PubMed
-
- Yatomi Y., Igarashi Y., Yang L., Hisano N., Qi R., Asazuma N., Satoh K., Ozaki Y., Kume S. (1997) Sphingosine 1-phosphate, a bioactive sphingolipid abundantly stored in platelets, is a normal constituent of human plasma and serum. J. Biochem. , 969–973 - PubMed
-
- Lin C. I., Chen C. N., Lin P. W., Lee H. (2007) Sphingosine 1-phosphate regulates inflammation-related genes in human endothelial cells through S1P1 and S1P3. Biochem. Biophys. Res. Commun. , 895–901 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
