

The Transmission Interfaces Contribute Asymmetrically to the Assembly and Activity of Human P-glycoprotein
- PMID: 25987565
- PMCID: PMC4505440
- DOI: 10.1074/jbc.M115.652602
The Transmission Interfaces Contribute Asymmetrically to the Assembly and Activity of Human P-glycoprotein
Abstract
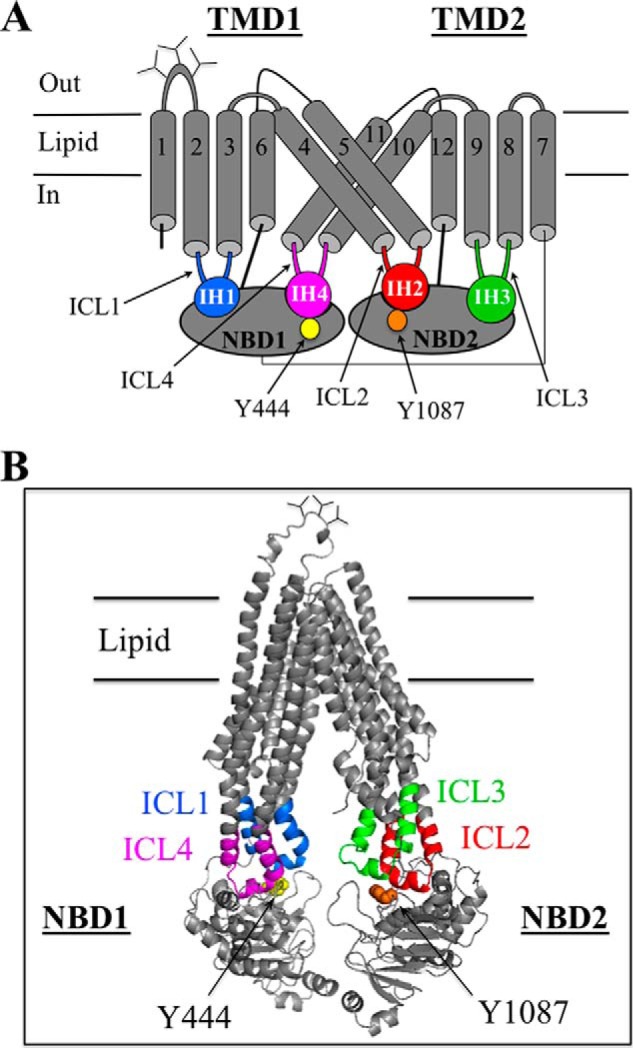
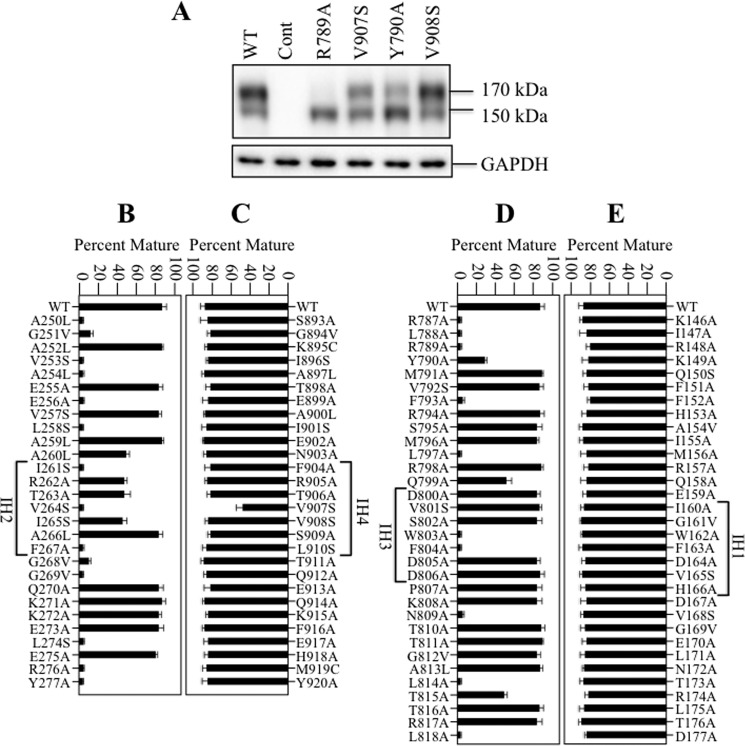
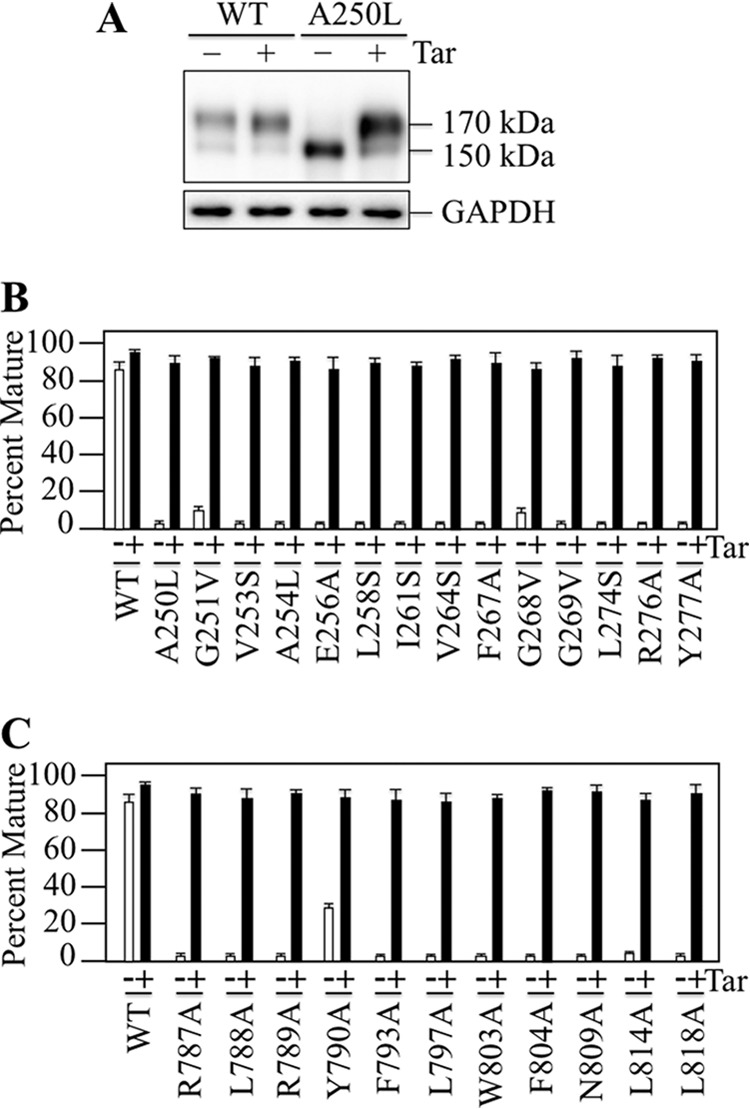
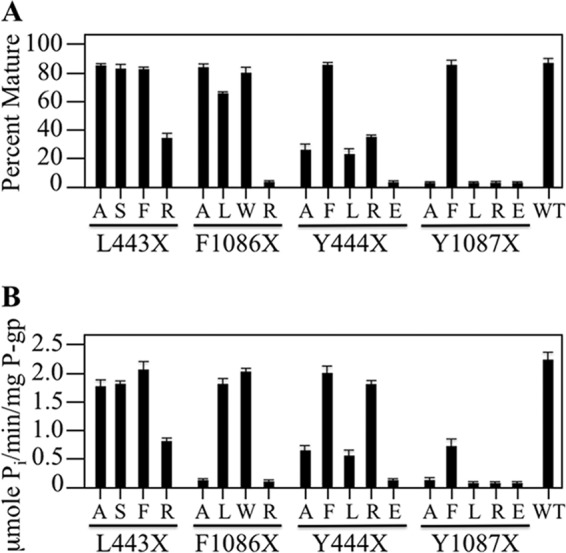
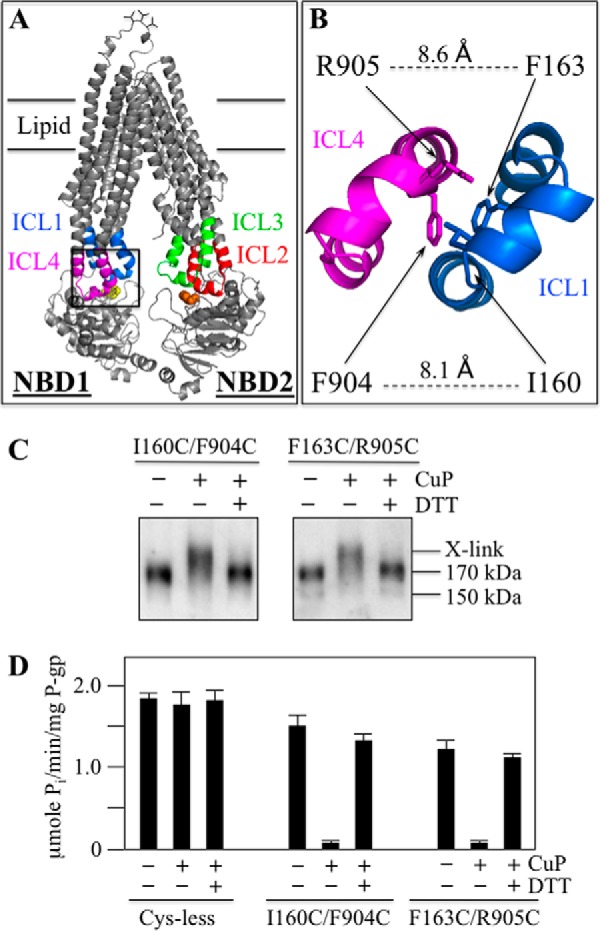
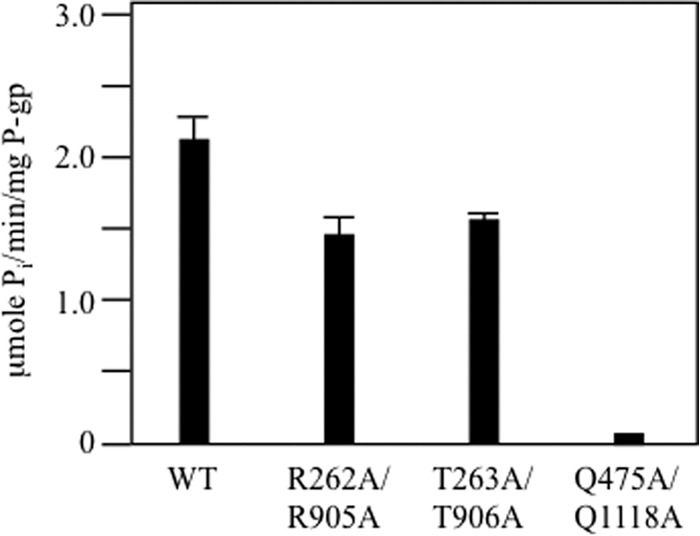
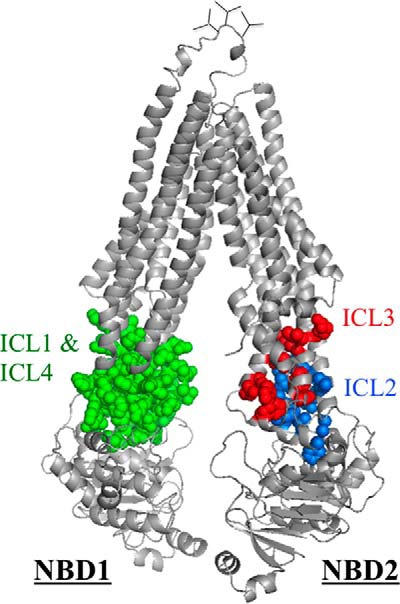
P-glycoprotein (P-gp; ABCB1) is an ABC drug pump that protects us from toxic compounds. It is clinically important because it confers multidrug resistance. The homologous halves of P-gp each contain a transmembrane (TM) domain (TMD) with 6 TM segments followed by a nucleotide-binding domain (NBD). The drug- and ATP-binding sites reside at the interface between the TMDs and NBDs, respectively. Each NBD is connected to the TMDs by a transmission interface involving a pair of intracellular loops (ICLs) that form ball-and-socket joints. P-gp is different from CFTR (ABCC7) in that deleting NBD2 causes misprocessing of only P-gp. Therefore, NBD2 might be critical for stabilizing ICLs 2 and 3 that form a tetrahelix bundle at the NBD2 interface. Here we report that the NBD1 and NBD2 transmission interfaces in P-gp are asymmetric. Point mutations to 25 of 60 ICL2/ICL3 residues at the NBD2 transmission interface severely reduced P-gp assembly while changes to the equivalent residues in ICL1/ICL4 at the NBD1 interface had little effect. The hydrophobic nature at the transmission interfaces was also different. Mutation of Phe-1086 or Tyr-1087 to arginine at the NBD2 socket blocked activity or assembly while the equivalent mutations at the NBD1 socket had only modest effects. The results suggest that the NBD transmission interfaces are asymmetric. In contrast to the ICL2/3-NBD2 interface, the ICL1/4-NBD1 transmission interface is more hydrophilic and insensitive to mutations. Therefore the ICL2/3-NBD2 transmission interface forms a precise hydrophobic connection that acts as a linchpin for assembly and trafficking of P-gp.
Keywords: ABC transporter; membrane enzyme; membrane protein; protein cross-linking; protein folding.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Borst P., Elferink R. O. (2002) Mammalian abc transporters in health and disease. Annu. Rev. Biochem. 71, 537–592 - PubMed
-
- Riordan J. R., Rommens J. M., Kerem B., Alon N., Rozmahel R., Grzelczak Z., Zielenski J., Lok S., Plavsic N., Chou J. L., Drumm M. L., Iannuzzi M. C., Collins F. S., Tsui L.-C. (1989) Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245, 1066–1073 - PubMed
-
- Nakagawa H., Toyoda Y., Wakabayashi-Nakao K., Tamaki H., Osumi M., Ishikawa T. (2011) Ubiquitin-mediated proteasomal degradation of ABC transporters: a new aspect of genetic polymorphisms and clinical impacts. J. Pharm. Sci. 100, 3602–3619 - PubMed
-
- Ostedgaard L. S., Rich D. P., DeBerg L. G., Welsh M. J. (1997) Association of domains within the cystic fibrosis transmembrane conductance regulator. Biochemistry 36, 1287–1294 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
