

Purinergic signalling during development and ageing
- PMID: 25989750
- PMCID: PMC4529855
- DOI: 10.1007/s11302-015-9452-9
Purinergic signalling during development and ageing
Abstract
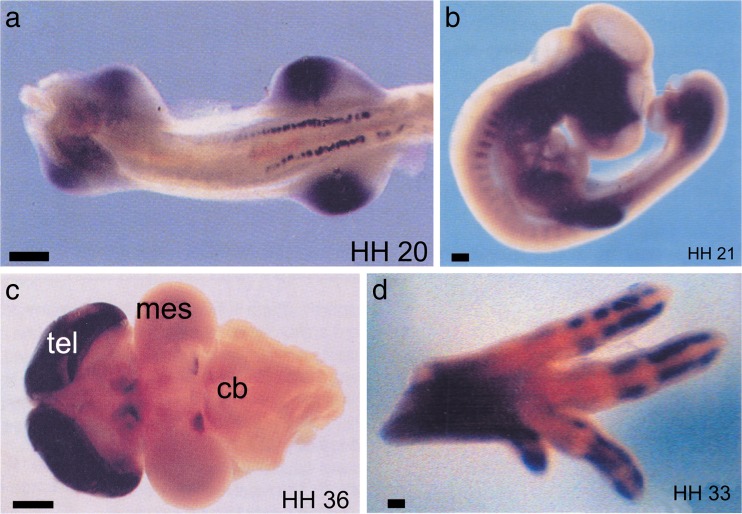
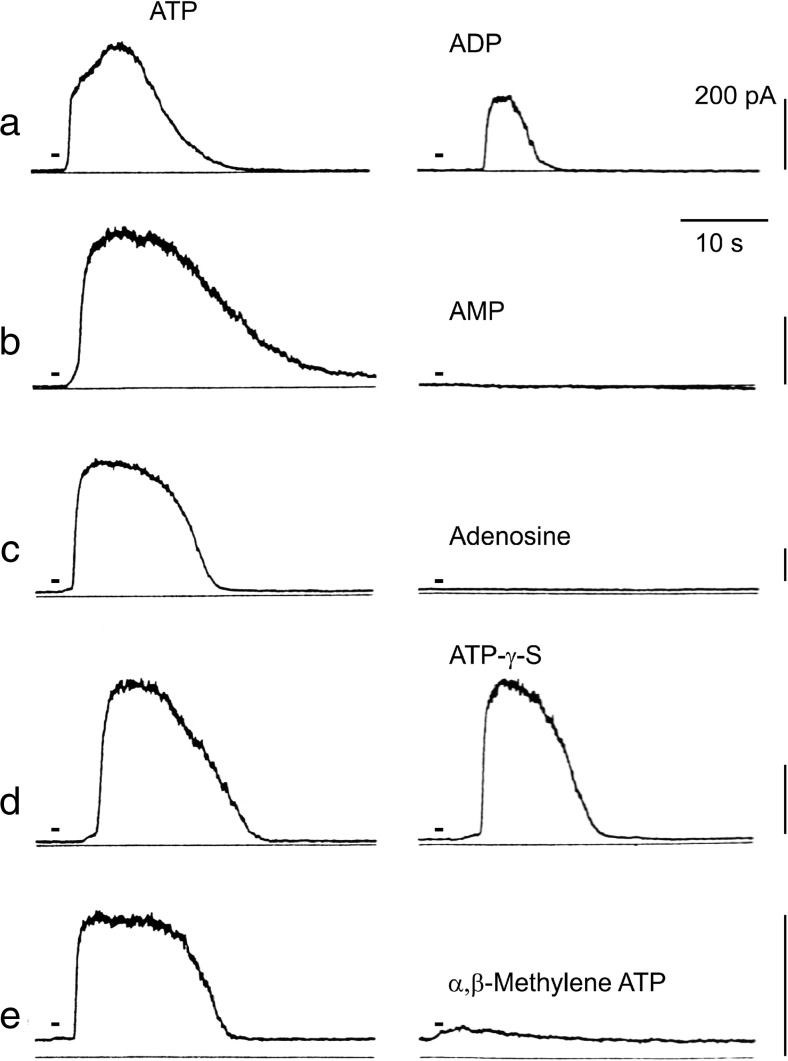
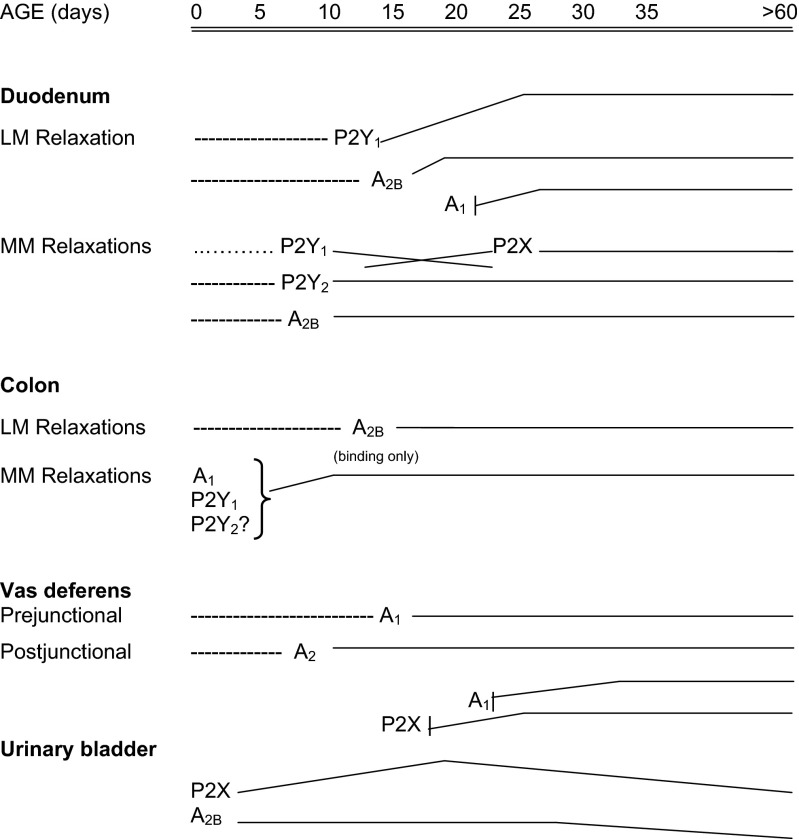
Extracellular purines and pyrimidines play major roles during embryogenesis, organogenesis, postnatal development and ageing in vertebrates, including humans. Pluripotent stem cells can differentiate into three primary germ layers of the embryo but may also be involved in plasticity and repair of the adult brain. These cells express the molecular components necessary for purinergic signalling, and their developmental fates can be manipulated via this signalling pathway. Functional P1, P2Y and P2X receptor subtypes and ectonucleotidases are involved in the development of different organ systems, including heart, blood vessels, skeletal muscle, urinary bladder, central and peripheral neurons, retina, inner ear, gut, lung and vas deferens. The importance of purinergic signalling in the ageing process is suggested by changes in expression of A1 and A2 receptors in old rat brains and reduction of P2X receptor expression in ageing mouse brain. By contrast, in the periphery, increases in expression of P2X3 and P2X4 receptors are seen in bladder and pancreas.
Figures
References
-
- Yeung CH. Temporary inhibition of the initiation of motility of demembranated hamster sperm by high concentrations of ATP. Int J Androl. 1986;9:359–370. - PubMed
-
- Foresta C, Rossato M, Di Virgilio F. Extracellular ATP is a trigger for the acrosome reaction in human spermatozoa. J Biol Chem. 1992;267:1–5. - PubMed
-
- Kupitz Y, Atlas D. A putative ATP-activated Na+ channel involved in sperm-induced fertilization. Science. 1993;261:484–486. - PubMed
-
- Smith R, Koenig C, Pereda J. Adenosinetriphosphatase (Mg-ATPase) activity in the plasma membrane of preimplantation mouse embryo as revealed by electron microscopy. Anat Embryol (Berl) 1983;168:455–466. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
