

The origin of snakes: revealing the ecology, behavior, and evolutionary history of early snakes using genomics, phenomics, and the fossil record
- PMID: 25989795
- PMCID: PMC4438441
- DOI: 10.1186/s12862-015-0358-5
The origin of snakes: revealing the ecology, behavior, and evolutionary history of early snakes using genomics, phenomics, and the fossil record
Abstract
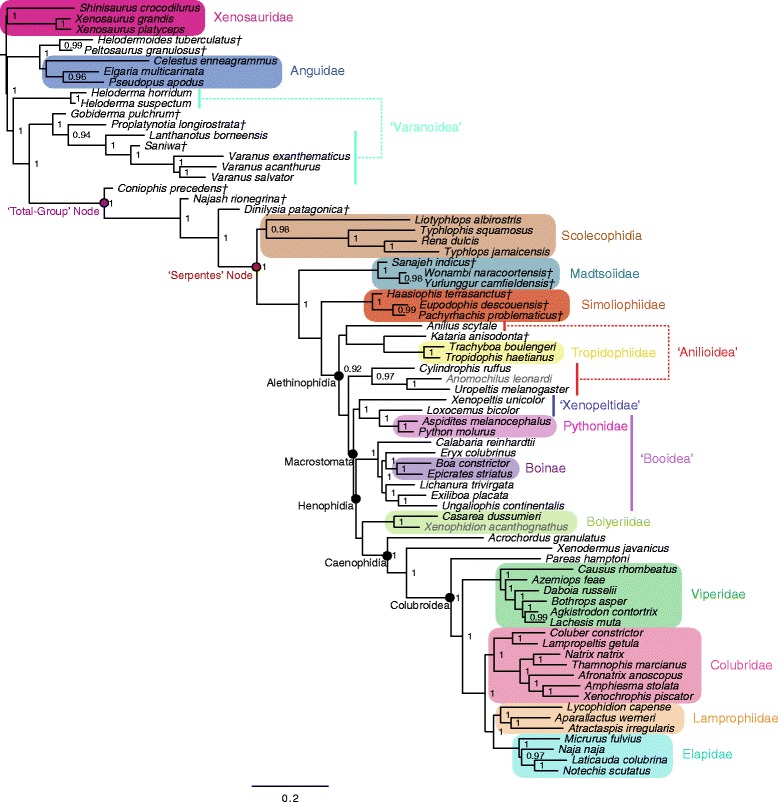
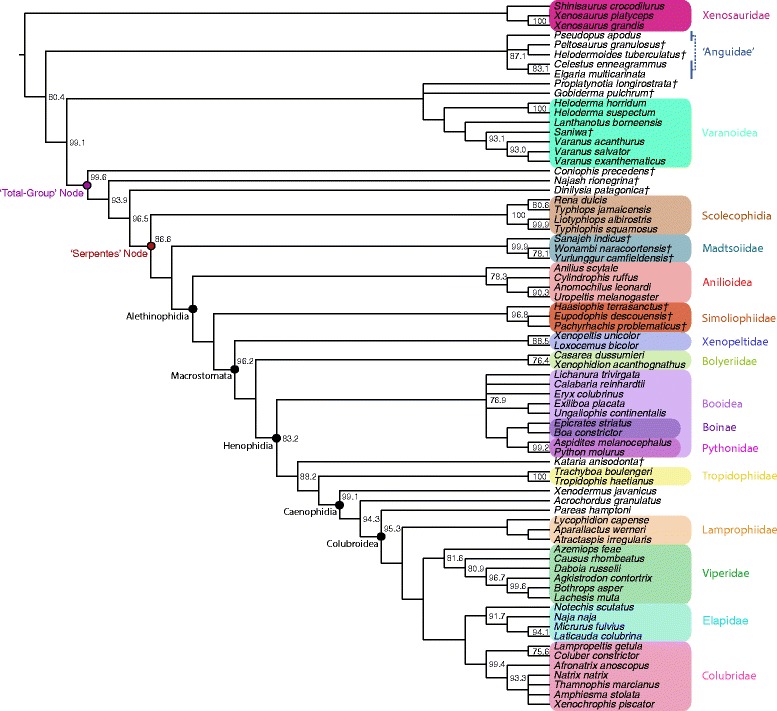
Background: The highly derived morphology and astounding diversity of snakes has long inspired debate regarding the ecological and evolutionary origin of both the snake total-group (Pan-Serpentes) and crown snakes (Serpentes). Although speculation abounds on the ecology, behavior, and provenance of the earliest snakes, a rigorous, clade-wide analysis of snake origins has yet to be attempted, in part due to a dearth of adequate paleontological data on early stem snakes. Here, we present the first comprehensive analytical reconstruction of the ancestor of crown snakes and the ancestor of the snake total-group, as inferred using multiple methods of ancestral state reconstruction. We use a combined-data approach that includes new information from the fossil record on extinct crown snakes, new data on the anatomy of the stem snakes Najash rionegrina, Dinilysia patagonica, and Coniophis precedens, and a deeper understanding of the distribution of phenotypic apomorphies among the major clades of fossil and Recent snakes. Additionally, we infer time-calibrated phylogenies using both new 'tip-dating' and traditional node-based approaches, providing new insights on temporal patterns in the early evolutionary history of snakes.
Results: Comprehensive ancestral state reconstructions reveal that both the ancestor of crown snakes and the ancestor of total-group snakes were nocturnal, widely foraging, non-constricting stealth hunters. They likely consumed soft-bodied vertebrate and invertebrate prey that was subequal to head size, and occupied terrestrial settings in warm, well-watered, and well-vegetated environments. The snake total-group - approximated by the Coniophis node - is inferred to have originated on land during the middle Early Cretaceous (~128.5 Ma), with the crown-group following about 20 million years later, during the Albian stage. Our inferred divergence dates provide strong evidence for a major radiation of henophidian snake diversity in the wake of the Cretaceous-Paleogene (K-Pg) mass extinction, clarifying the pattern and timing of the extant snake radiation. Although the snake crown-group most likely arose on the supercontinent of Gondwana, our results suggest the possibility that the snake total-group originated on Laurasia.
Conclusions: Our study provides new insights into when, where, and how snakes originated, and presents the most complete picture of the early evolution of snakes to date. More broadly, we demonstrate the striking influence of including fossils and phenotypic data in combined analyses aimed at both phylogenetic topology inference and ancestral state reconstruction.
Figures







References
-
- Scanlon JD, Lee MSY. The major clades of living snakes. In: Aldridge RB, Sever DM, editors. Reproductive Biology and Phylogeny of Snakes. Enfield, New Hampshire: Science Publishers, Inc.; 2011. pp. 55–95.
-
- Webb JK, Shine R, Branch WR, Harlow PS. Life-history strategy in basal snakes: reproduction and dietary habits of the African thread snake Leptotyphlops scutifrons (Serpentes: Leptotyphlopidae) J Zool Lond. 2000;250:321–7. doi: 10.1111/j.1469-7998.2000.tb00776.x. - DOI
-
- Zaher H, Scanferla CA. The skull of the Upper Cretaceous snake Dinilysia patagonica Smith-Woodward, 1901, and its phylogenetic position revisited. Zool J Linn Soc. 2012;164:194–238. doi: 10.1111/j.1096-3642.2011.00755.x. - DOI
-
- Zaher H, Apesteguía S, Scanferla CA. The anatomy of the upper cretaceous snake Najash rionegrina Apesteguía & Zaher, 2006, and the evolution of limblessness in snakes. Zool J Linn Soc. 2009;156:801–26. doi: 10.1111/j.1096-3642.2009.00511.x. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
