

Quantitative Analysis of Glutamate Receptors in Glial Cells from the Cortex of GFAP/EGFP Mice Following Ischemic Injury: Focus on NMDA Receptors
- PMID: 25994914
- PMCID: PMC11486180
- DOI: 10.1007/s10571-015-0212-8
Quantitative Analysis of Glutamate Receptors in Glial Cells from the Cortex of GFAP/EGFP Mice Following Ischemic Injury: Focus on NMDA Receptors
Abstract
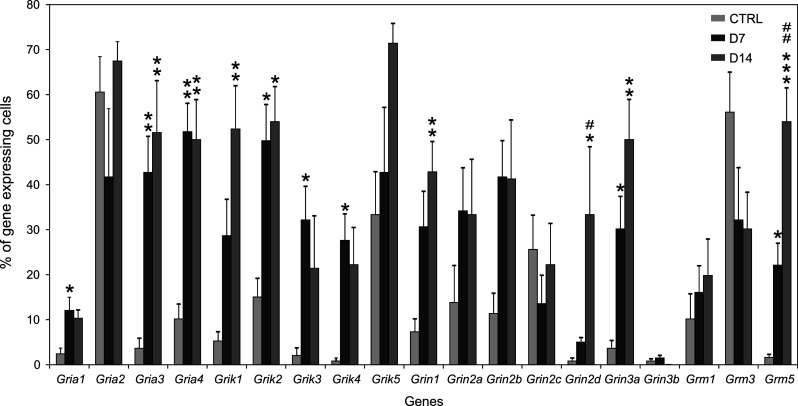
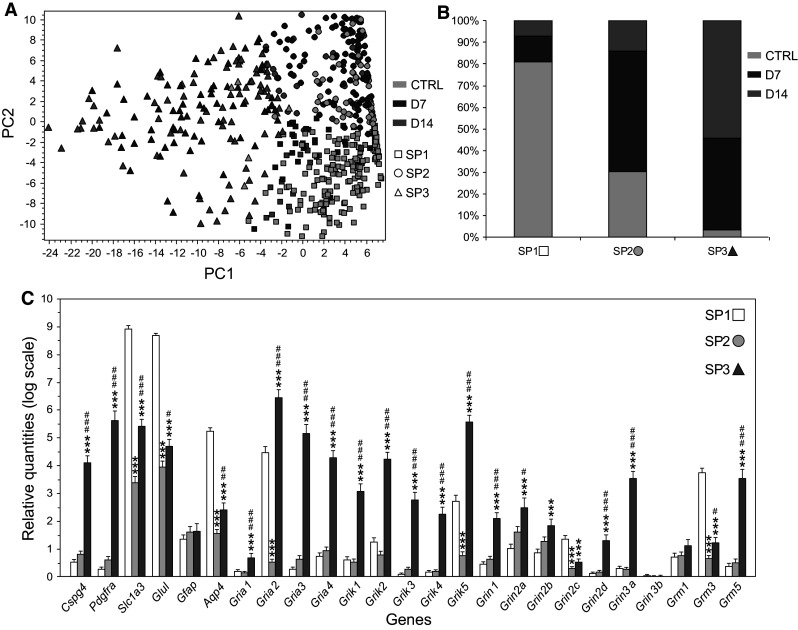
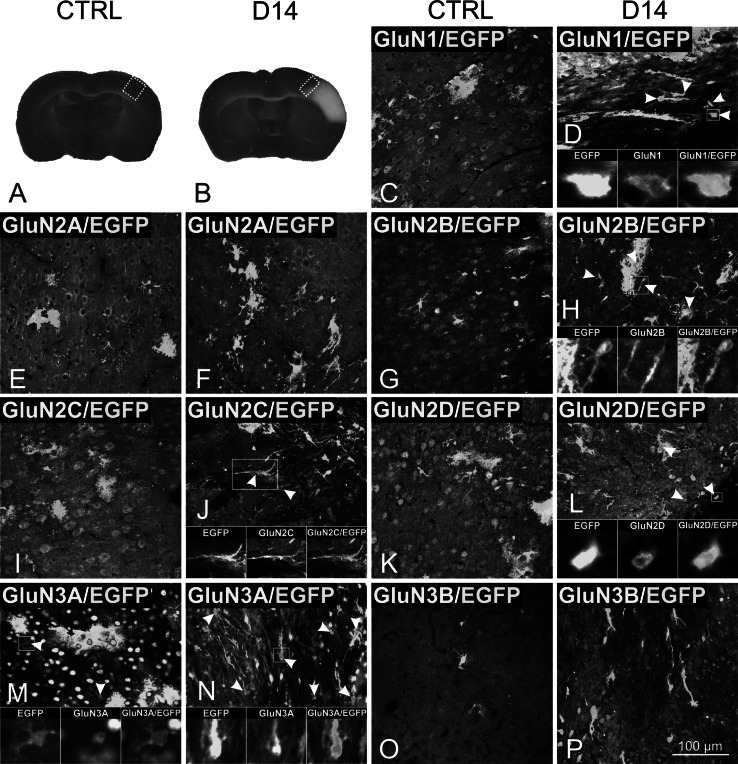
Cortical glial cells contain both ionotropic and metabotropic glutamate receptors. Despite several efforts, a comprehensive analysis of the entire family of glutamate receptors and their subunits present in glial cells is still missing. Here, we provide an overall picture of the gene expression of ionotropic (AMPA, kainate, NMDA) and the main metabotropic glutamate receptors in cortical glial cells isolated from GFAP/EGFP mice before and after focal cerebral ischemia. Employing single-cell RT-qPCR, we detected the expression of genes encoding subunits of glutamate receptors in GFAP/EGFP-positive (GFAP/EGFP(+)) glial cells in the cortex of young adult mice. Most of the analyzed cells expressed mRNA for glutamate receptor subunits, the expression of which, in most cases, even increased after ischemic injury. Data analyses disclosed several classes of GFAP/EGFP(+) glial cells with respect to glutamate receptors and revealed in what manner their expression correlates with the expression of glial markers prior to and after ischemia. Furthermore, we also examined the protein expression and functional significance of NMDA receptors in glial cells. Immunohistochemical analyses of all seven NMDA receptor subunits provided direct evidence that the GluN3A subunit is present in GFAP/EGFP(+) glial cells and that its expression is increased after ischemia. In situ and in vitro Ca(2+) imaging revealed that Ca(2+) elevations evoked by the application of NMDA were diminished in GFAP/EGFP(+) glial cells following ischemia. Our results provide a comprehensive description of glutamate receptors in cortical GFAP/EGFP(+) glial cells and may serve as a basis for further research on glial cell physiology and pathophysiology.
Keywords: Astrocytes; Calcium imaging; MCAo; NG2 glia; Single-cell RT-qPCR.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Aronica E, van Vliet EA, Mayboroda OA et al (2000) Upregulation of metabotropic glutamate receptor subtype mGluR3 and mGluR5 in reactive astrocytes in a rat model of mesial temporal lobe epilepsy. Eur J Neurosci 12:2333–2344 - PubMed
-
- Aronica E, Leenstra S, Jansen GH et al (2001) Expression of brain-derived neurotrophic factor and tyrosine kinase B receptor proteins in glioneuronal tumors from patients with intractable epilepsy: colocalization with N-methyl-D-aspartic acid receptor. Acta Neuropathol 101:383–392 - PubMed
-
- Aronica E, Gorter JA, Ijlst-Keizers H et al (2003) Expression and functional role of mGluR3 and mGluR5 in human astrocytes and glioma cells: opposite regulation of glutamate transporter proteins. Eur J Neurosci 17:2106–2118. doi:10.1046/j.1460-9568.2003.02657.x - PubMed
-
- Atlason PT, Garside ML, Meddows E et al (2007) N-Methyl-D-aspartate (NMDA) receptor subunit NR1 forms the substrate for oligomeric assembly of the NMDA receptor. J Biol Chem 282:25299–25307. doi:10.1074/jbc.M702778200 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
