

P-body assembly requires DDX6 repression complexes rather than decay or Ataxin2/2L complexes
- PMID: 25995375
- PMCID: PMC4501357
- DOI: 10.1091/mbc.E15-03-0136
P-body assembly requires DDX6 repression complexes rather than decay or Ataxin2/2L complexes
Abstract
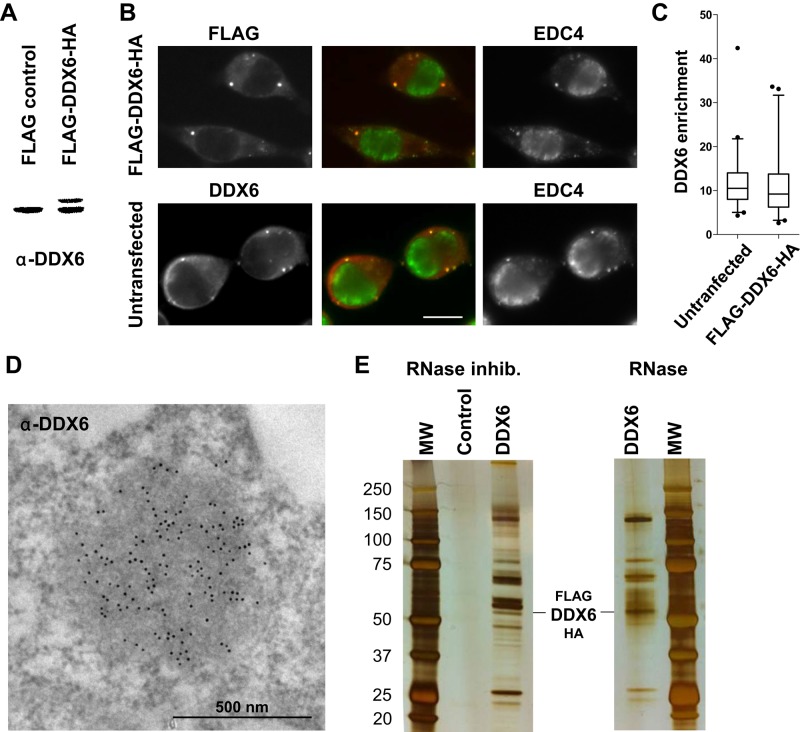
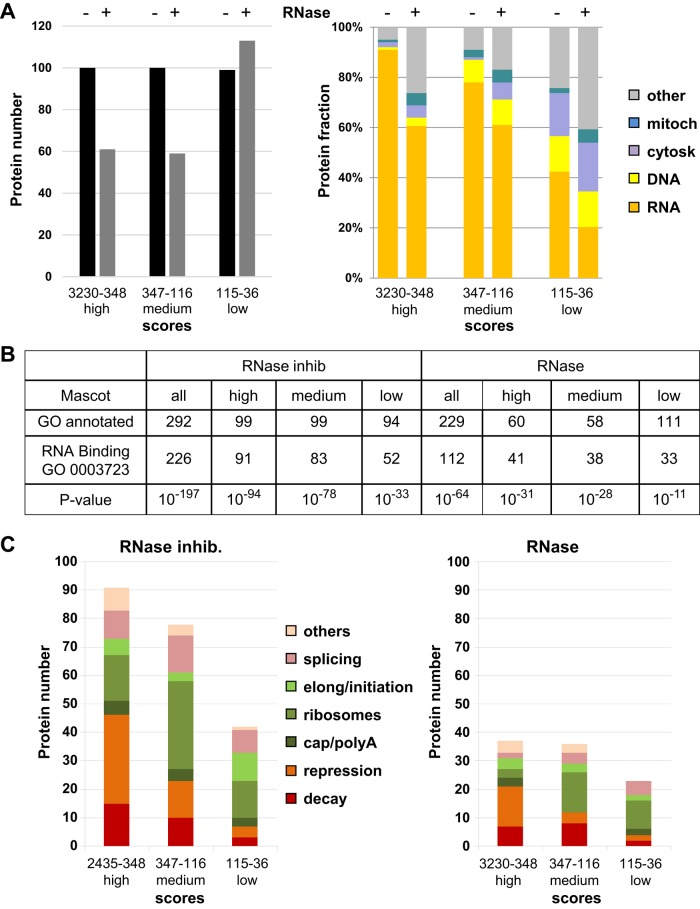
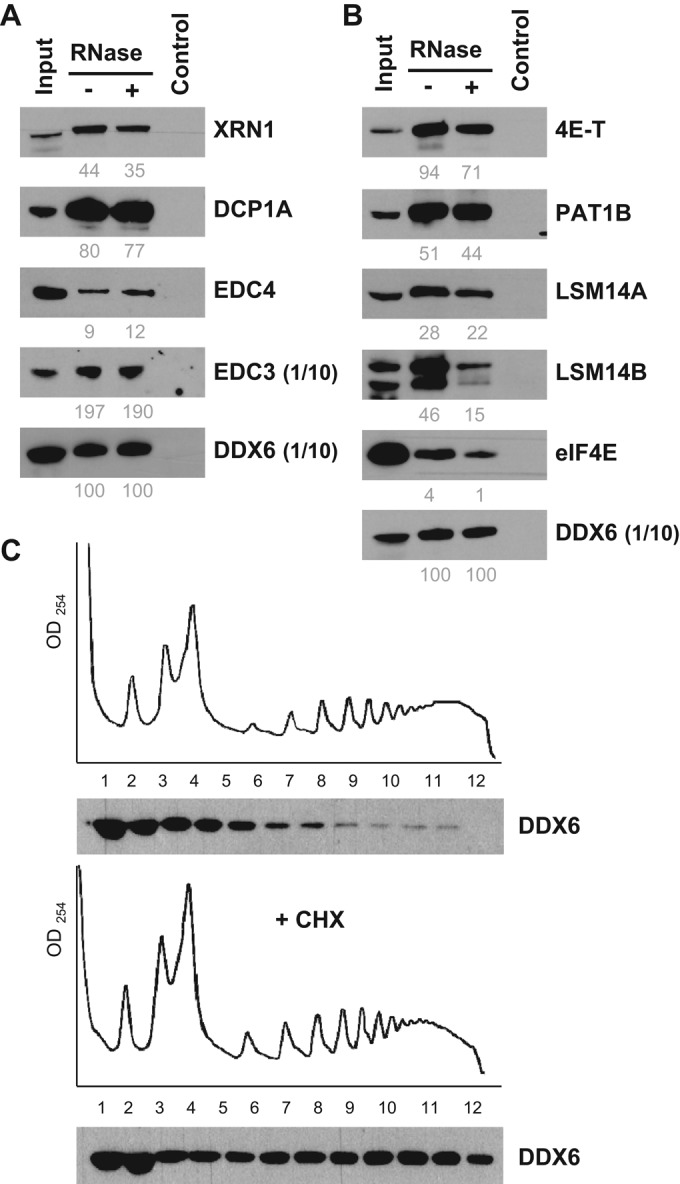
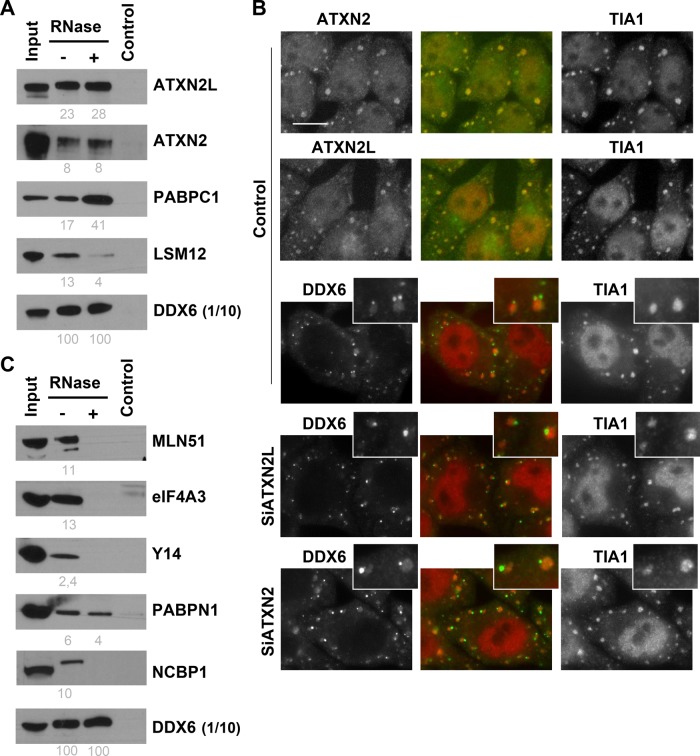
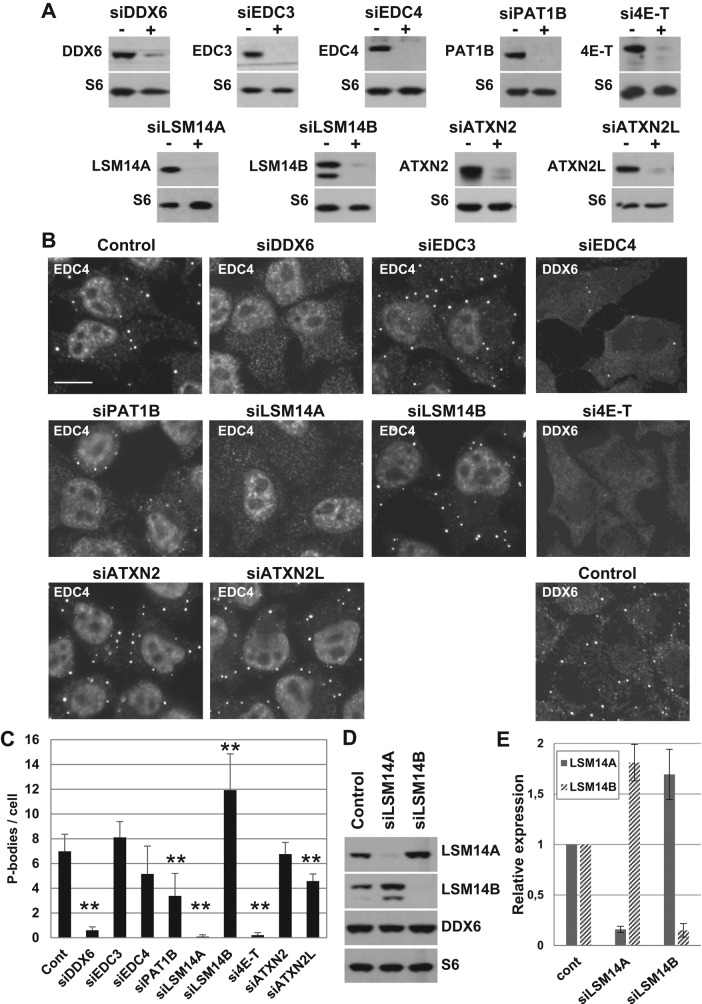
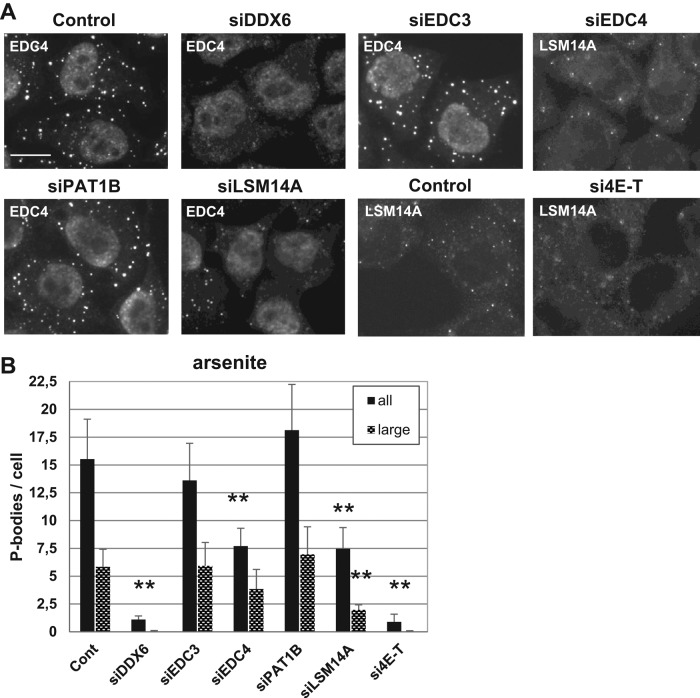
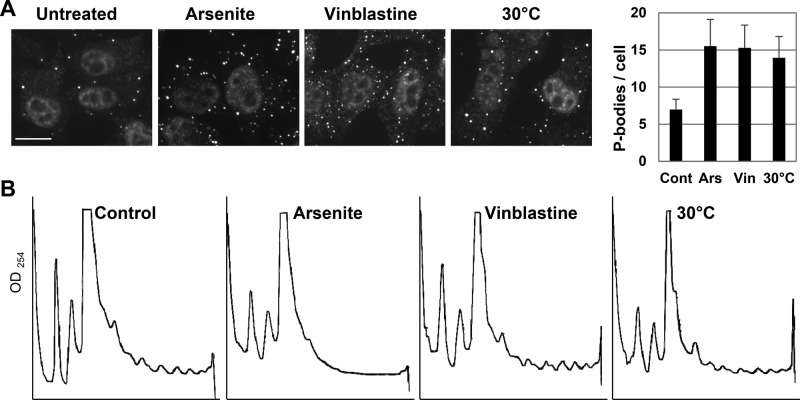
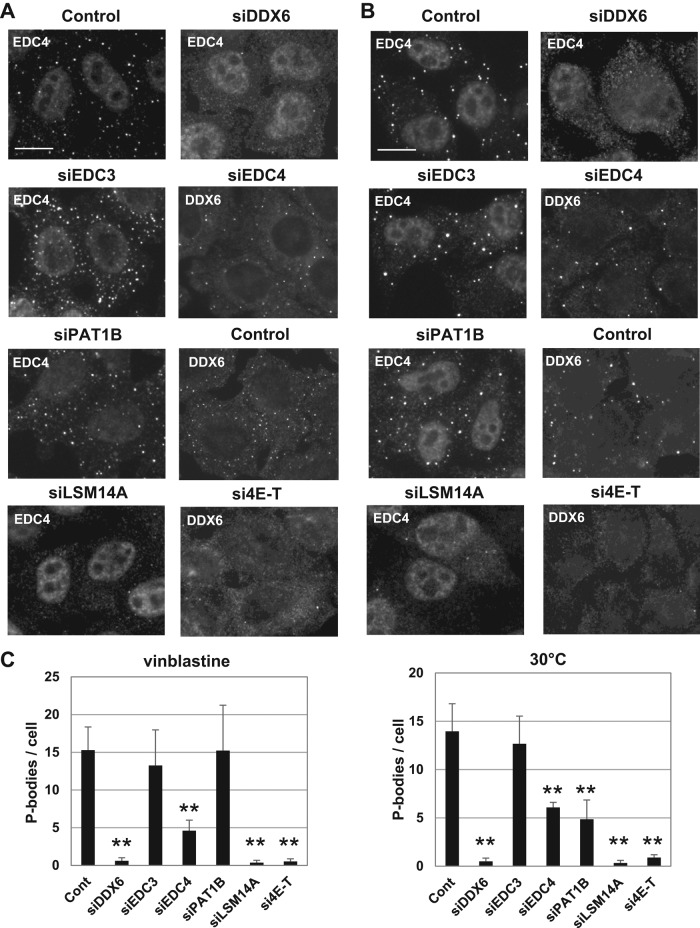
P-bodies are cytoplasmic ribonucleoprotein granules involved in posttranscriptional regulation. DDX6 is a key component of their assembly in human cells. This DEAD-box RNA helicase is known to be associated with various complexes, including the decapping complex, the CPEB repression complex, RISC, and the CCR4/NOT complex. To understand which DDX6 complexes are required for P-body assembly, we analyzed the DDX6 interactome using the tandem-affinity purification methodology coupled to mass spectrometry. Three complexes were prominent: the decapping complex, a CPEB-like complex, and an Ataxin2/Ataxin2L complex. The exon junction complex was also found, suggesting DDX6 binding to newly exported mRNAs. Finally, some DDX6 was associated with polysomes, as previously reported in yeast. Despite its high enrichment in P-bodies, most DDX6 is localized out of P-bodies. Of the three complexes, only the decapping and CPEB-like complexes were recruited into P-bodies. Investigation of P-body assembly in various conditions allowed us to distinguish required proteins from those that are dispensable or participate only in specific conditions. Three proteins were required in all tested conditions: DDX6, 4E-T, and LSM14A. These results reveal the variety of pathways of P-body assembly, which all nevertheless share three key factors connecting P-body assembly to repression.
© 2015 Ayache, Bénard, et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Chen Y, Boland A, Kuzuog˘lu-Öztürk D, Bawankar P, Loh B, Chang C-T, Weichenrieder O, Izaurralde E. A DDX6-CNOT1 complex and W-binding pockets in CNOT9 reveal direct links between miRNA target recognition and silencing. Mol Cell. 2014;54:737–750. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
