

Characterization of Novel Ribosome-Associated Endoribonuclease SLFN14 from Rabbit Reticulocytes
- PMID: 25996083
- PMCID: PMC4461289
- DOI: 10.1021/acs.biochem.5b00302
Characterization of Novel Ribosome-Associated Endoribonuclease SLFN14 from Rabbit Reticulocytes
Abstract
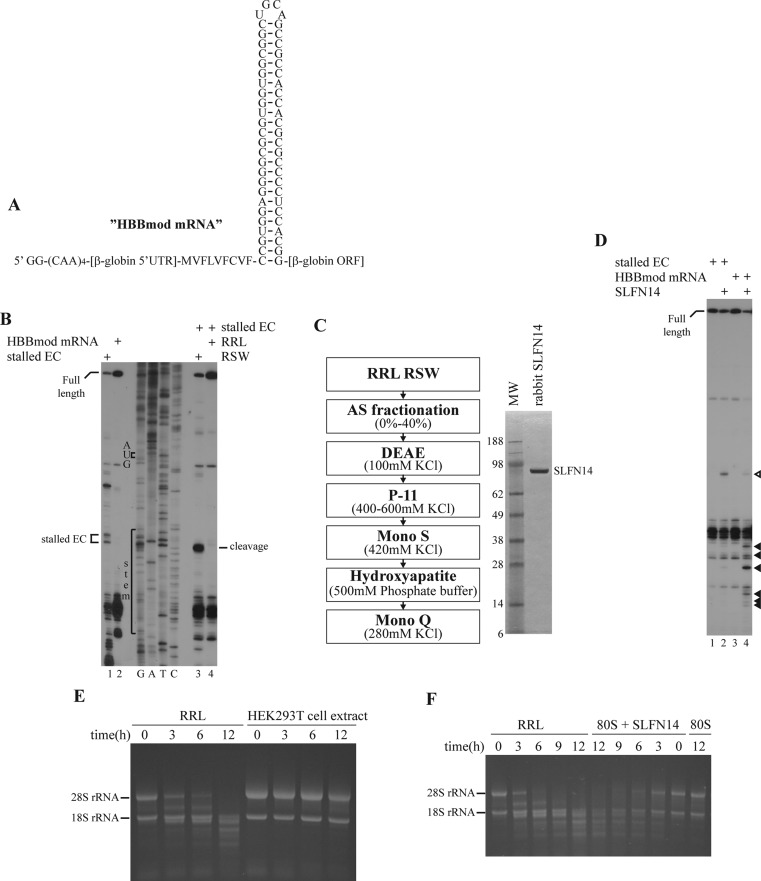
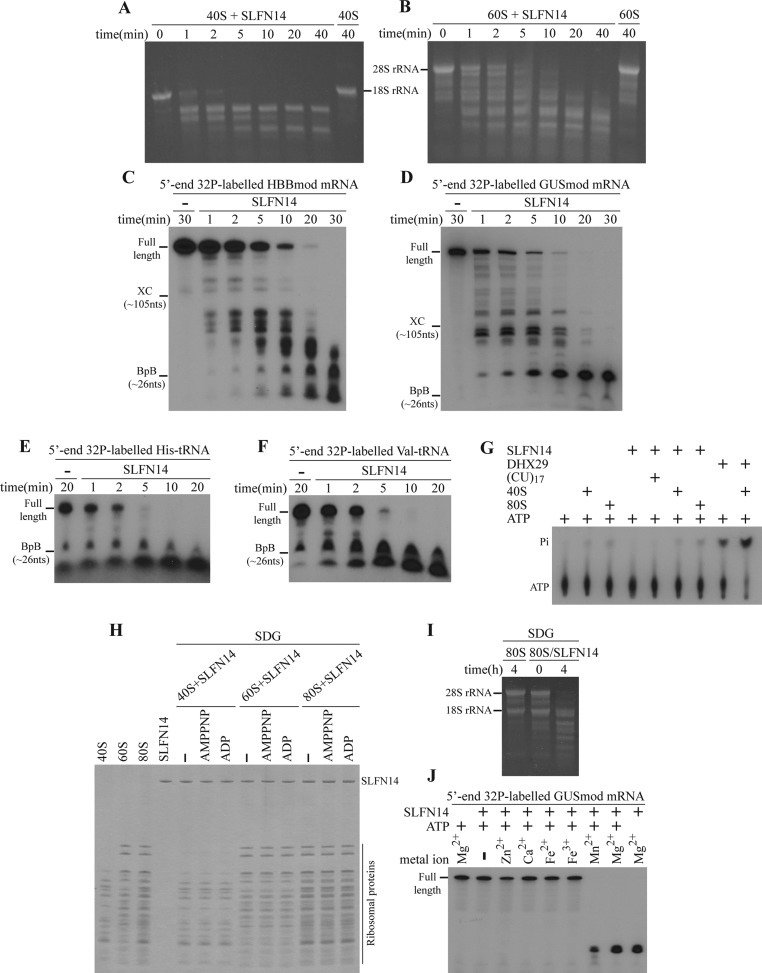
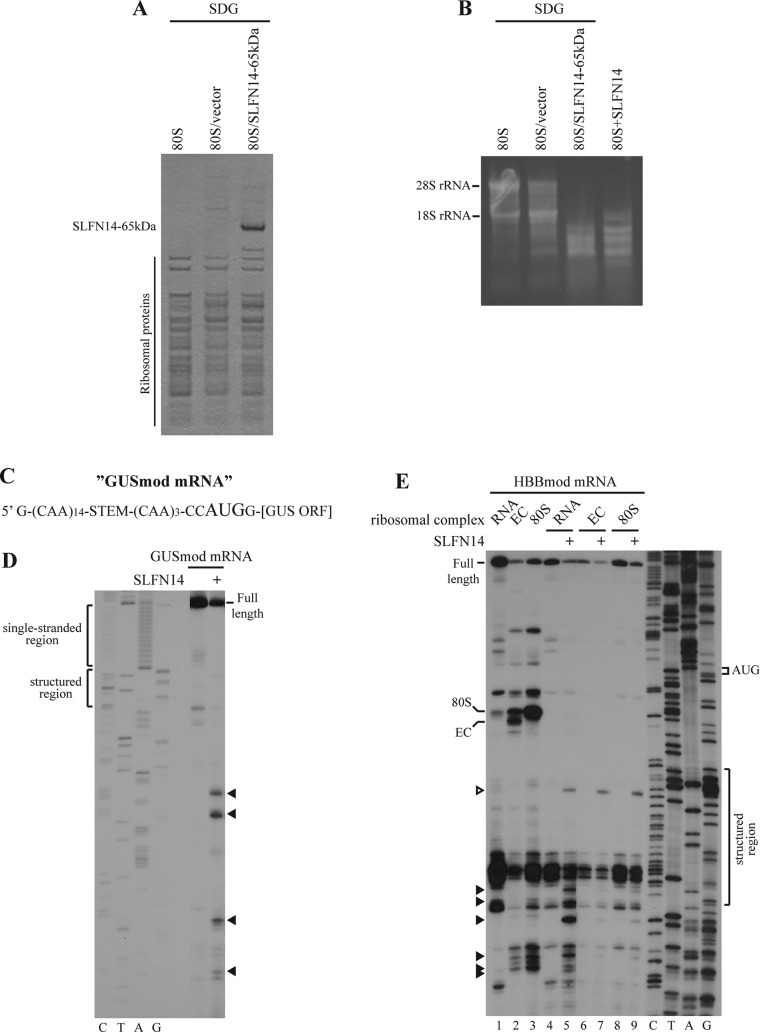
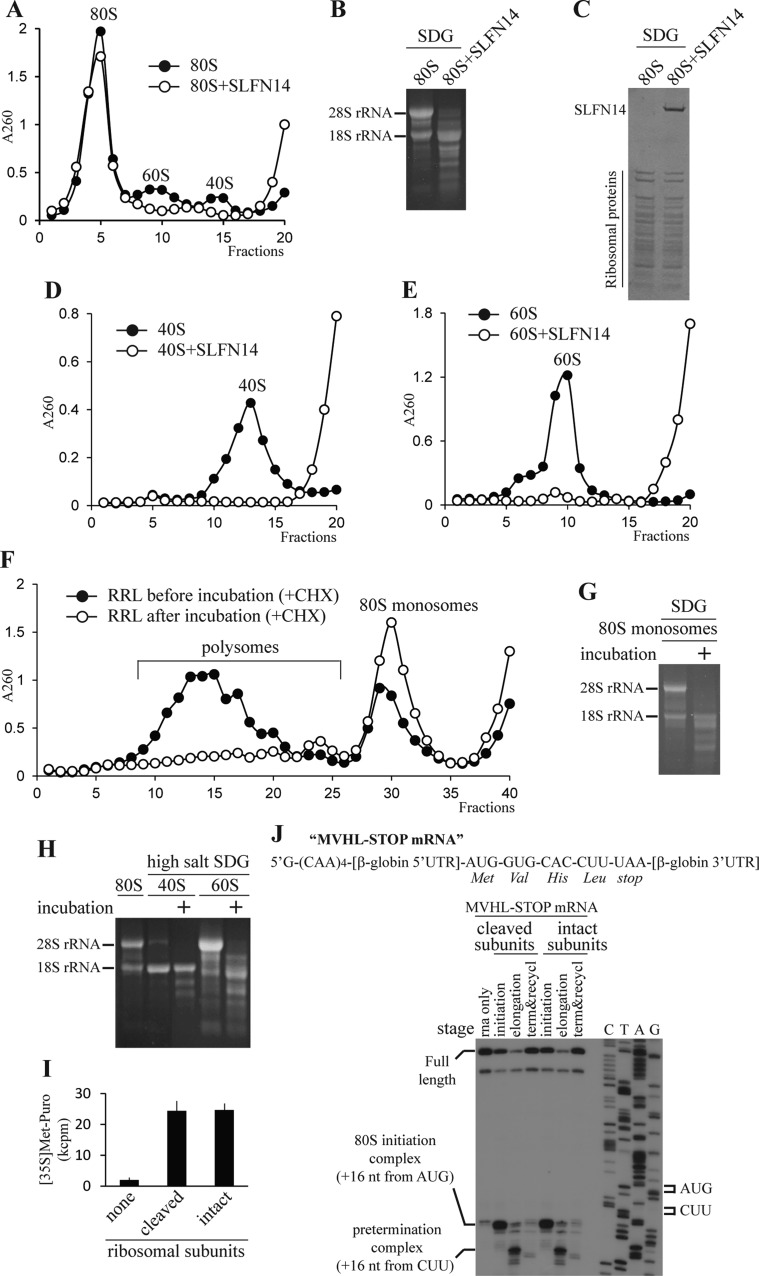
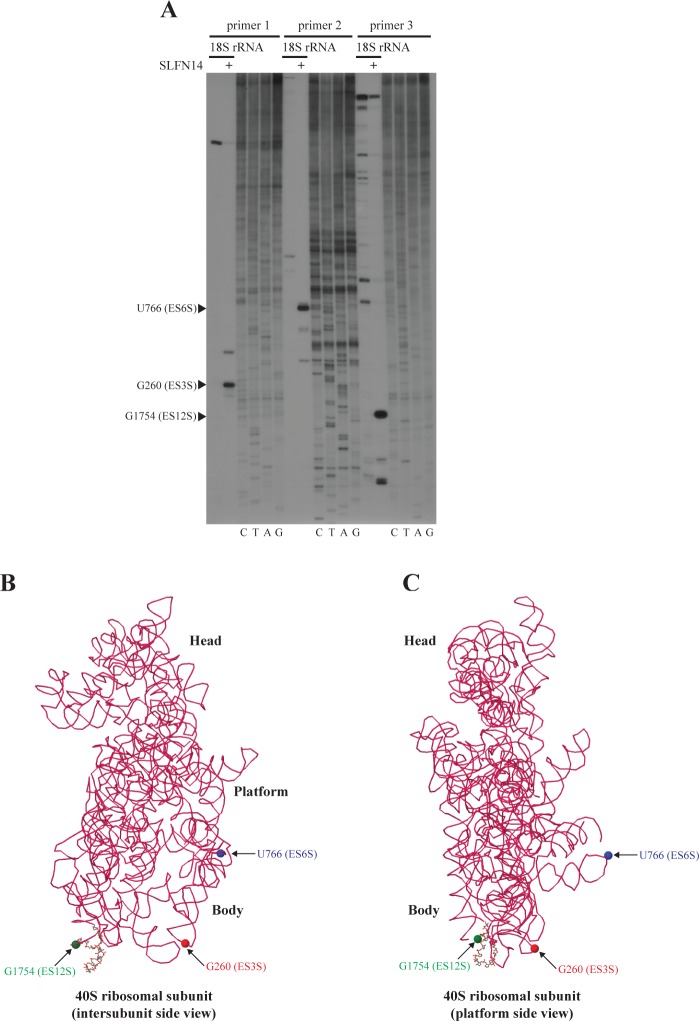
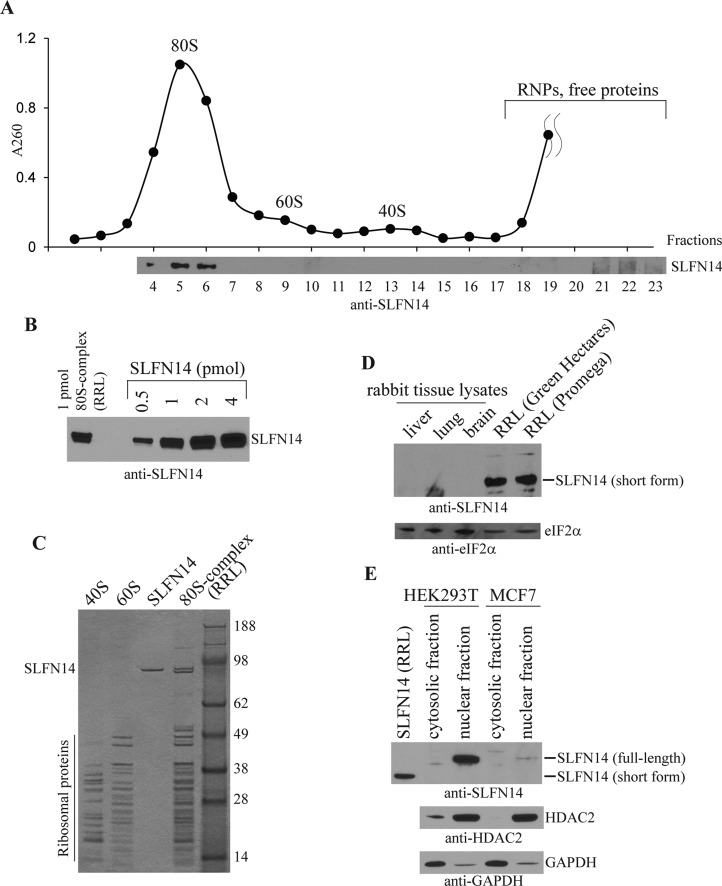
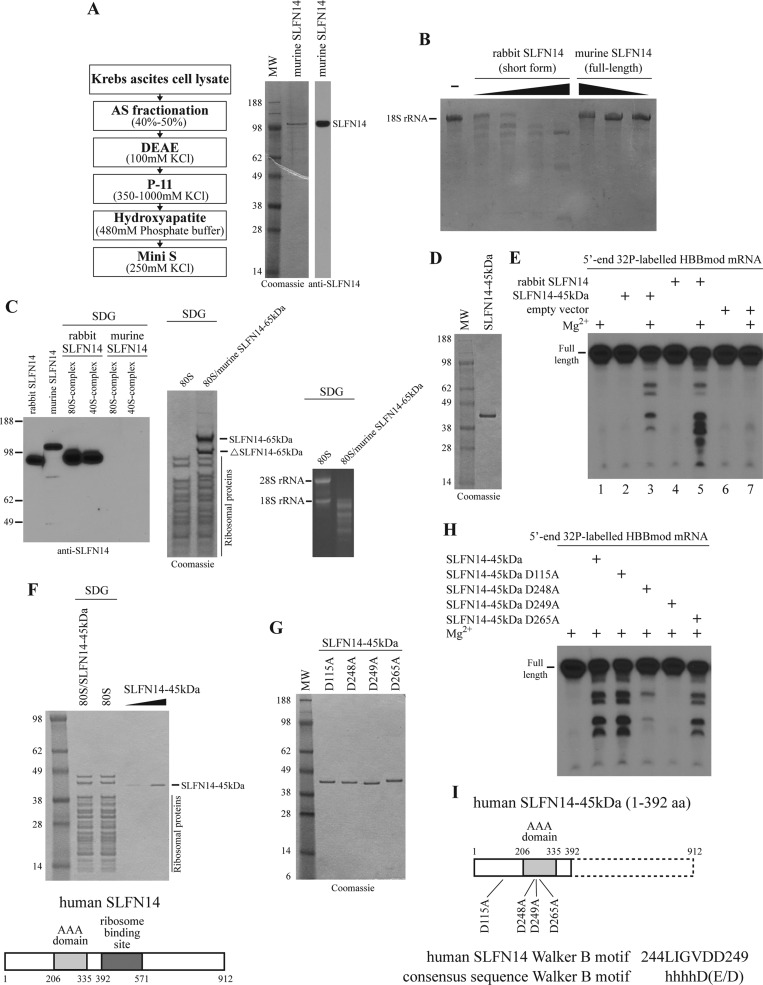
Turnover of mRNA is a critical step that allows cells to control gene expression. Endoribonucleases, enzymes cleaving RNA molecules internally, are some of the key components of the degradation process. Here we provide a detailed characterization of novel endoribonuclease SLFN14 purified from rabbit reticulocyte lysate. Schlafen genes encode a family of proteins limited to mammals. Their cellular function is unknown or incompletely understood. In reticulocytes, SLFN14 is strongly overexpressed, represented exclusively by the short form, all tethered to ribosomes, and appears to be one of the major ribosome-associated proteins. SLFN14 binds to ribosomes and ribosomal subunits in the low part of the body and cleaves RNA but preferentially rRNA and ribosome-associated mRNA. This results in the degradation of ribosomal subunits. This process is strictly Mg(2+)- and Mn(2+)-dependent, NTP-independent, and sequence nonspecific. However, in other cell types, SLFN14 is a full-length solely nuclear protein, which lacks ribosomal binding and nuclease activities. Mutational analysis revealed the ribosomal binding site and the aspartate essential for the endonucleolytic activity of protein. Only few endoribonucleases participating in ribosome-mediated processes have been characterized to date. Moreover, none of them are shown to be directly associated with the ribosome. Therefore, our findings expand the general knowledge of endoribonucleases involved in mammalian translation control.
Figures
References
-
- Li W. M.; Barnes T.; Lee C. H. (2010) Endoribonucleases: Enzymes gaining spotlight in mRNA metabolism. FEBS J. 277, 627–641. - PubMed
-
- Walter P.; Ron D. (2011) The unfolded protein response: From stress pathway to homeostatic regulation. Science 334, 1081–1086. - PubMed
-
- Hollien J.; Weissman J. S. (2006) Decay of endoplasmic reticulum-localized mRNAs during the unfolded protein response. Science 313, 104–107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
