

Turning Escherichia coli into a Frataxin-Dependent Organism
- PMID: 25996492
- PMCID: PMC4440780
- DOI: 10.1371/journal.pgen.1005134
Turning Escherichia coli into a Frataxin-Dependent Organism
Abstract
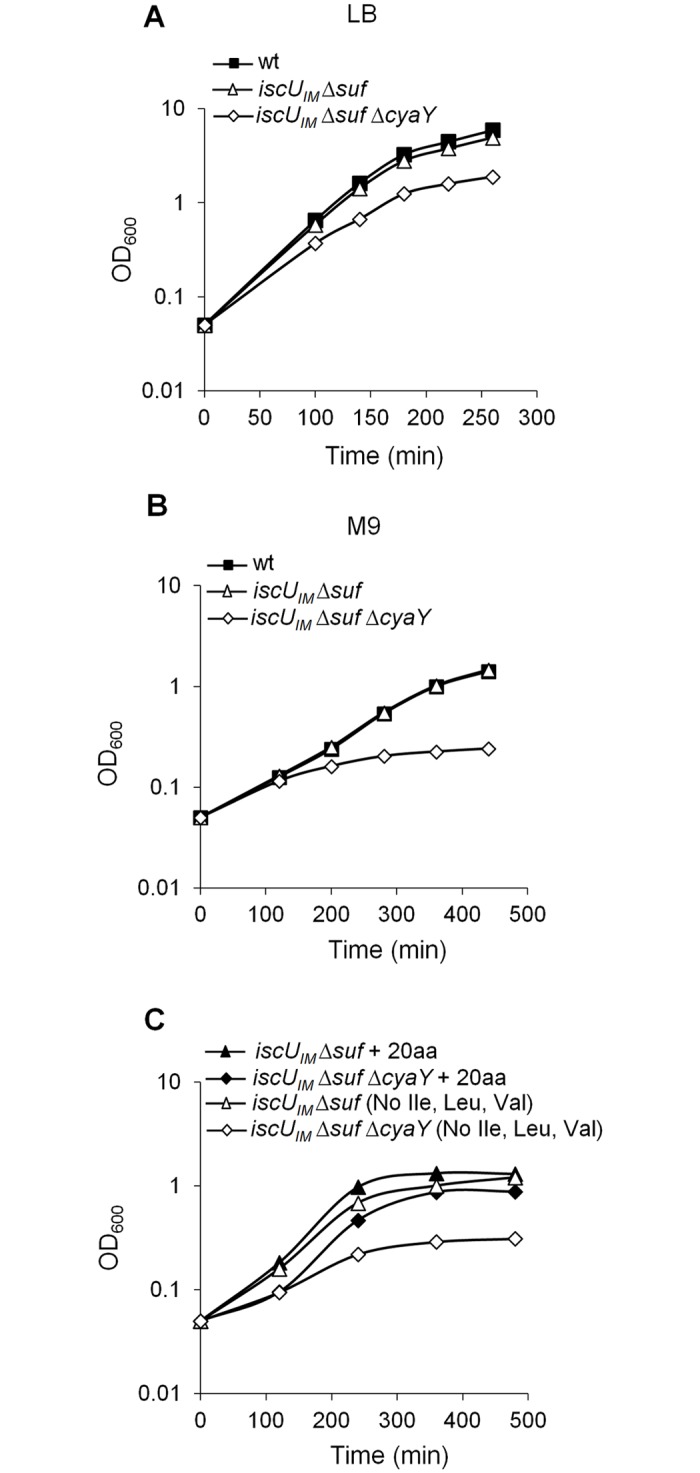
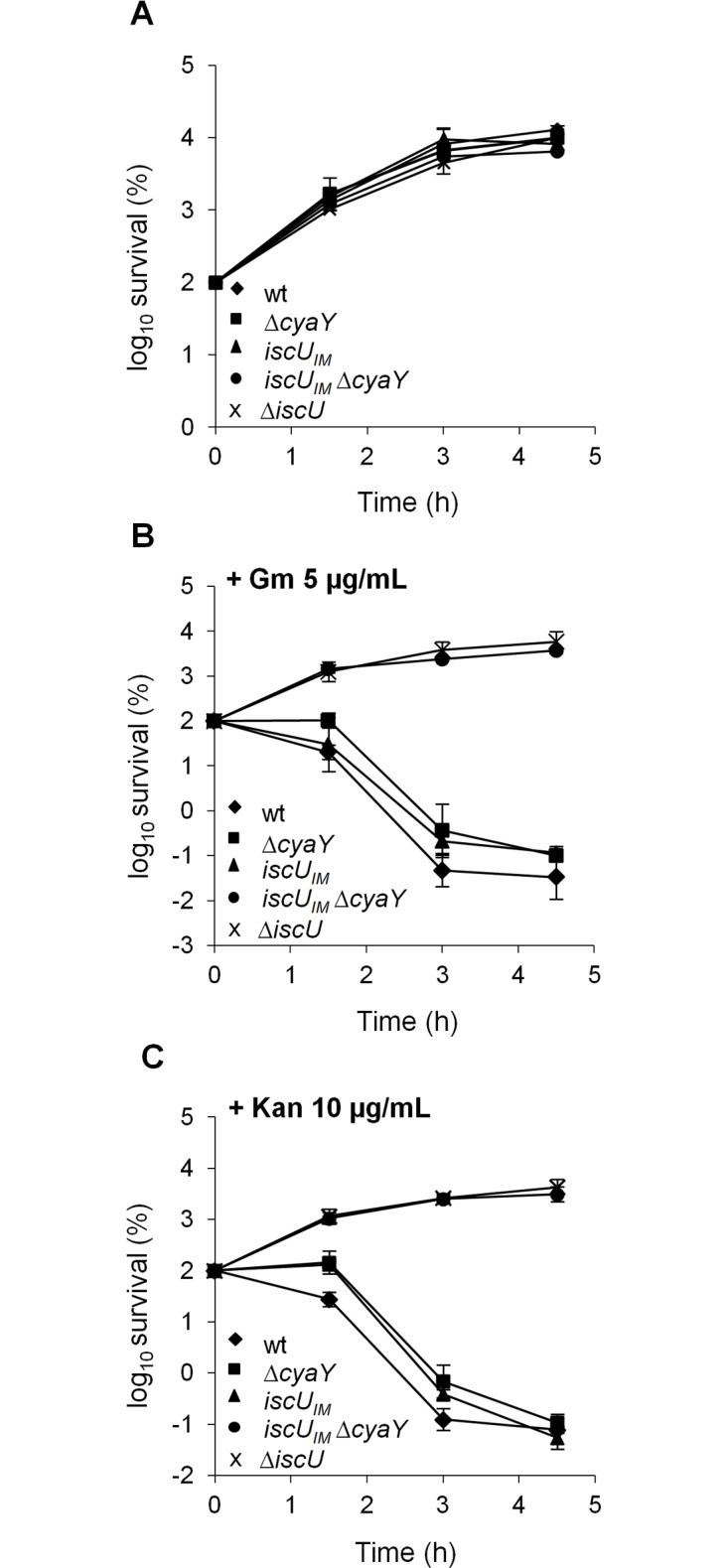
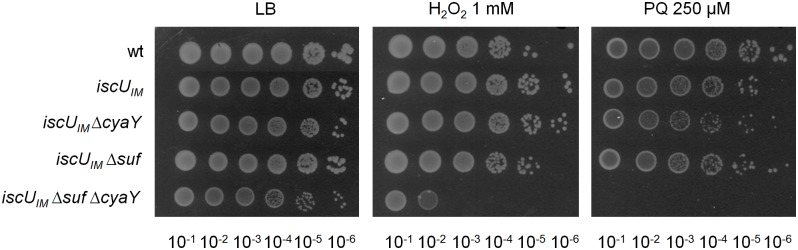
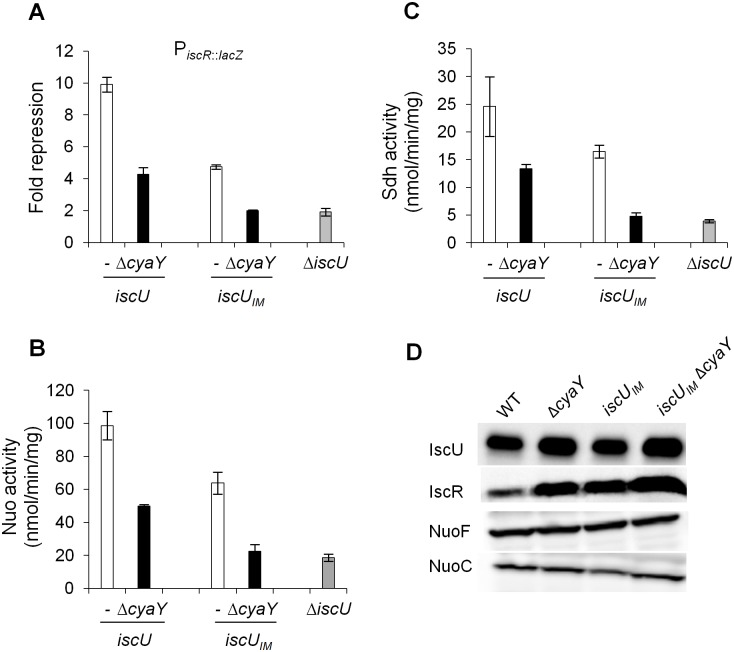
Fe-S bound proteins are ubiquitous and contribute to most basic cellular processes. A defect in the ISC components catalyzing Fe-S cluster biogenesis leads to drastic phenotypes in both eukaryotes and prokaryotes. In this context, the Frataxin protein (FXN) stands out as an exception. In eukaryotes, a defect in FXN results in severe defects in Fe-S cluster biogenesis, and in humans, this is associated with Friedreich's ataxia, a neurodegenerative disease. In contrast, prokaryotes deficient in the FXN homolog CyaY are fully viable, despite the clear involvement of CyaY in ISC-catalyzed Fe-S cluster formation. The molecular basis of the differing importance in the contribution of FXN remains enigmatic. Here, we have demonstrated that a single mutation in the scaffold protein IscU rendered E. coli viability strictly dependent upon a functional CyaY. Remarkably, this mutation changed an Ile residue, conserved in prokaryotes at position 108, into a Met residue, conserved in eukaryotes. We found that in the double mutant IscUIM ΔcyaY, the ISC pathway was completely abolished, becoming equivalent to the ΔiscU deletion strain and recapitulating the drastic phenotype caused by FXN deletion in eukaryotes. Biochemical analyses of the "eukaryotic-like" IscUIM scaffold revealed that it exhibited a reduced capacity to form Fe-S clusters. Finally, bioinformatic studies of prokaryotic IscU proteins allowed us to trace back the source of FXN-dependency as it occurs in present-day eukaryotes. We propose an evolutionary scenario in which the current mitochondrial Isu proteins originated from the IscUIM version present in the ancestor of the Rickettsiae. Subsequent acquisition of SUF, the second Fe-S cluster biogenesis system, in bacteria, was accompanied by diminished contribution of CyaY in prokaryotic Fe-S cluster biogenesis, and increased tolerance to change in the amino acid present at the 108th position of the scaffold.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
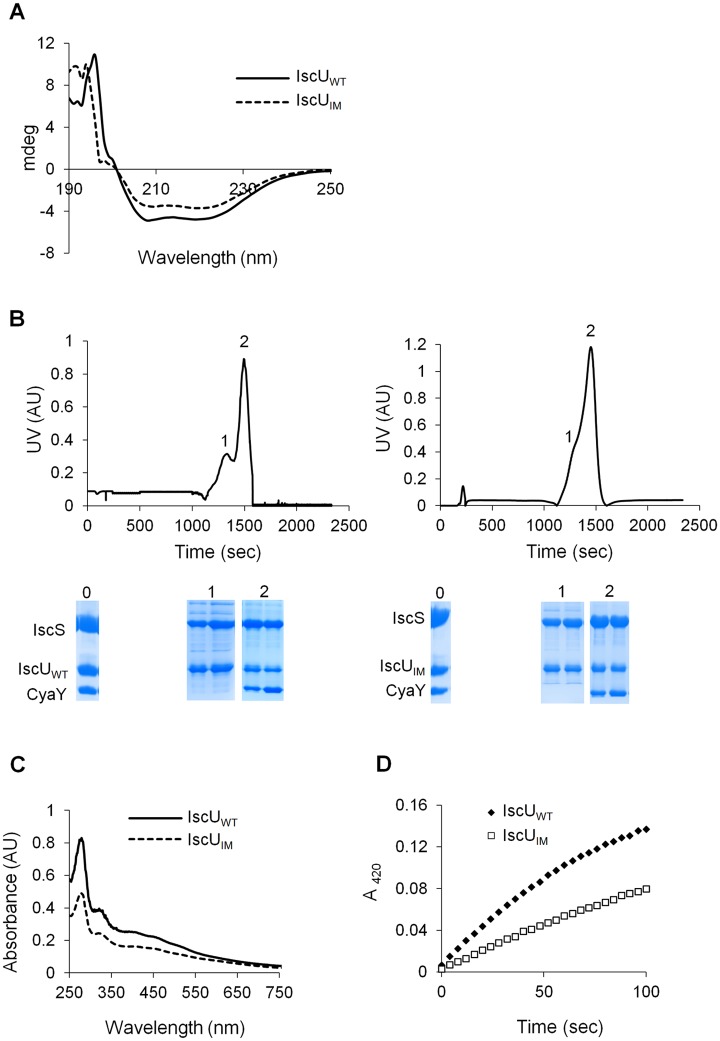
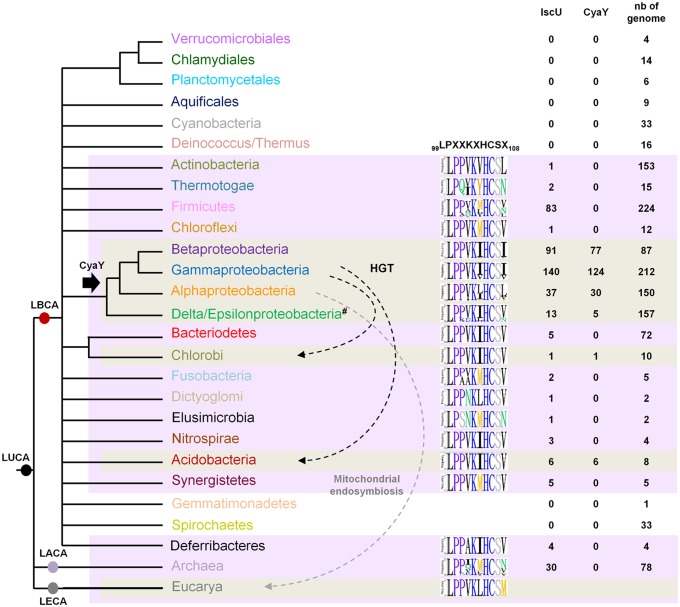
Comment in
-
Trading Places-Switching Frataxin Function by a Single Amino Acid Substitution within the [Fe-S] Cluster Assembly Scaffold.PLoS Genet. 2015 May 21;11(5):e1005192. doi: 10.1371/journal.pgen.1005192. eCollection 2015 May. PLoS Genet. 2015. PMID: 25996679 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
