

The kinase DYRK1A reciprocally regulates the differentiation of Th17 and regulatory T cells
- PMID: 25998054
- PMCID: PMC4441007
- DOI: 10.7554/eLife.05920
The kinase DYRK1A reciprocally regulates the differentiation of Th17 and regulatory T cells
Abstract
The balance between Th17 and T regulatory (Treg) cells critically modulates immune homeostasis, with an inadequate Treg response contributing to inflammatory disease. Using an unbiased chemical biology approach, we identified a novel role for the dual specificity tyrosine-phosphorylation-regulated kinase DYRK1A in regulating this balance. Inhibition of DYRK1A enhances Treg differentiation and impairs Th17 differentiation without affecting known pathways of Treg/Th17 differentiation. Thus, DYRK1A represents a novel mechanistic node at the branch point between commitment to either Treg or Th17 lineages. Importantly, both Treg cells generated using the DYRK1A inhibitor harmine and direct administration of harmine itself potently attenuate inflammation in multiple experimental models of systemic autoimmunity and mucosal inflammation. Our results identify DYRK1A as a physiologically relevant regulator of Treg cell differentiation and suggest a broader role for other DYRK family members in immune homeostasis. These results are discussed in the context of human diseases associated with dysregulated DYRK activity.
Keywords: T cell differentiation; computational biology; dual-specificity tyrosine-regulated kinase signaling; human; immunology; inflammation; mouse; systems biology.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
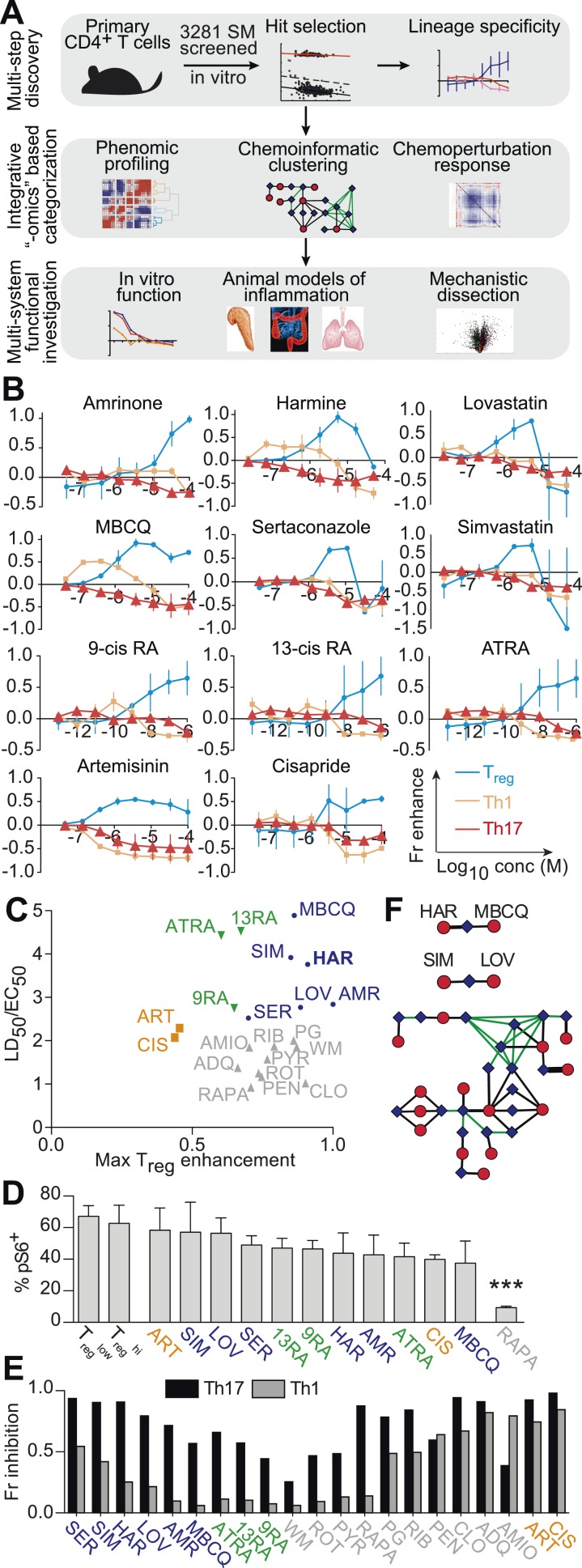
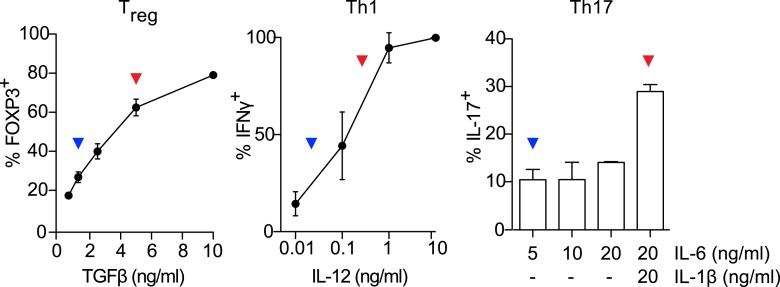
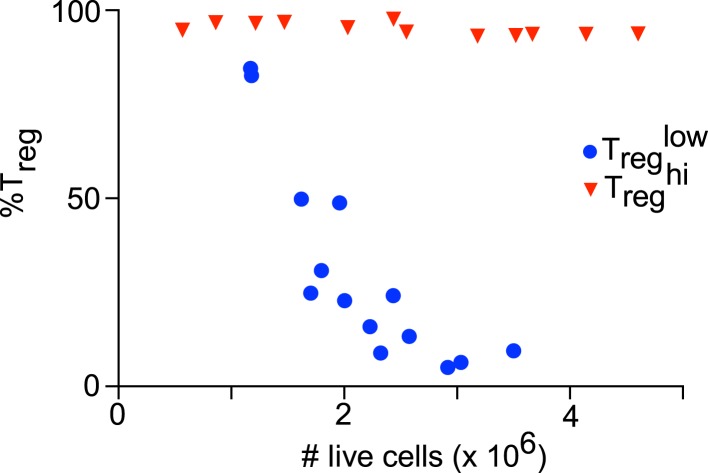
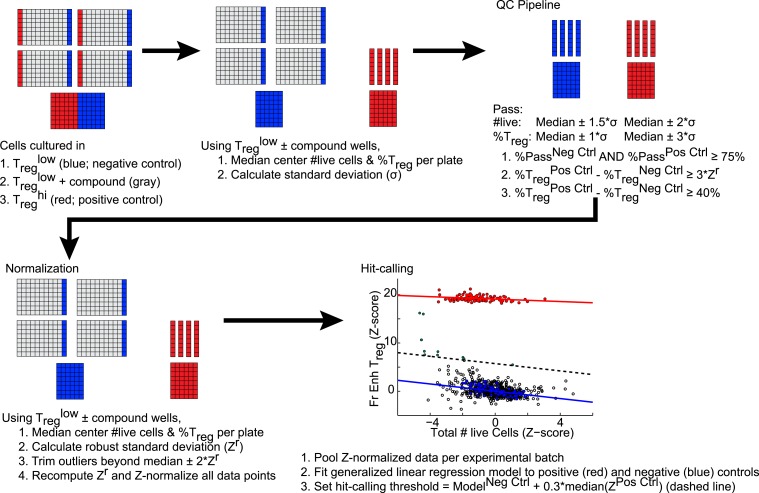
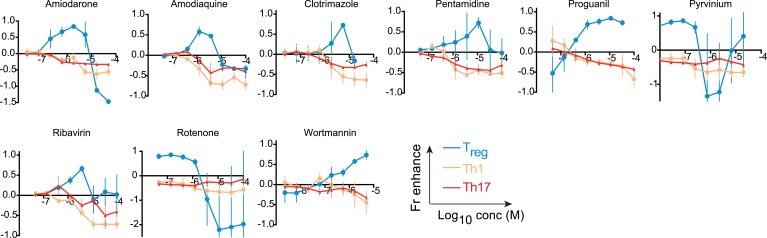
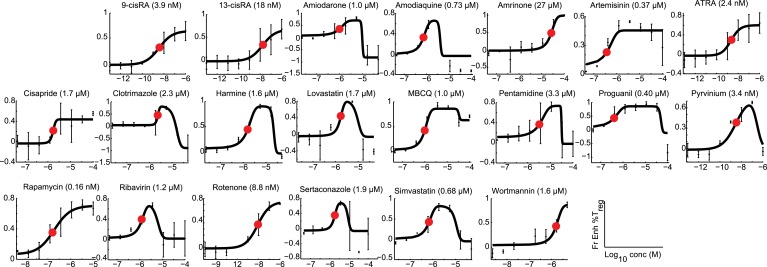
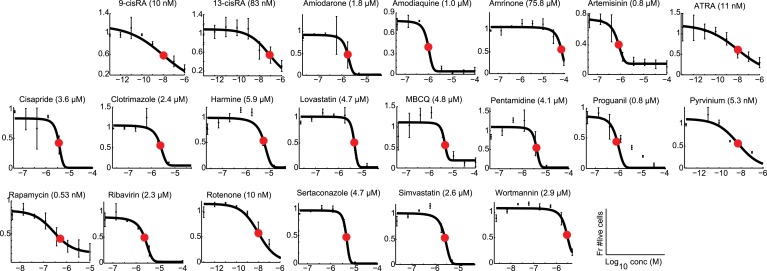
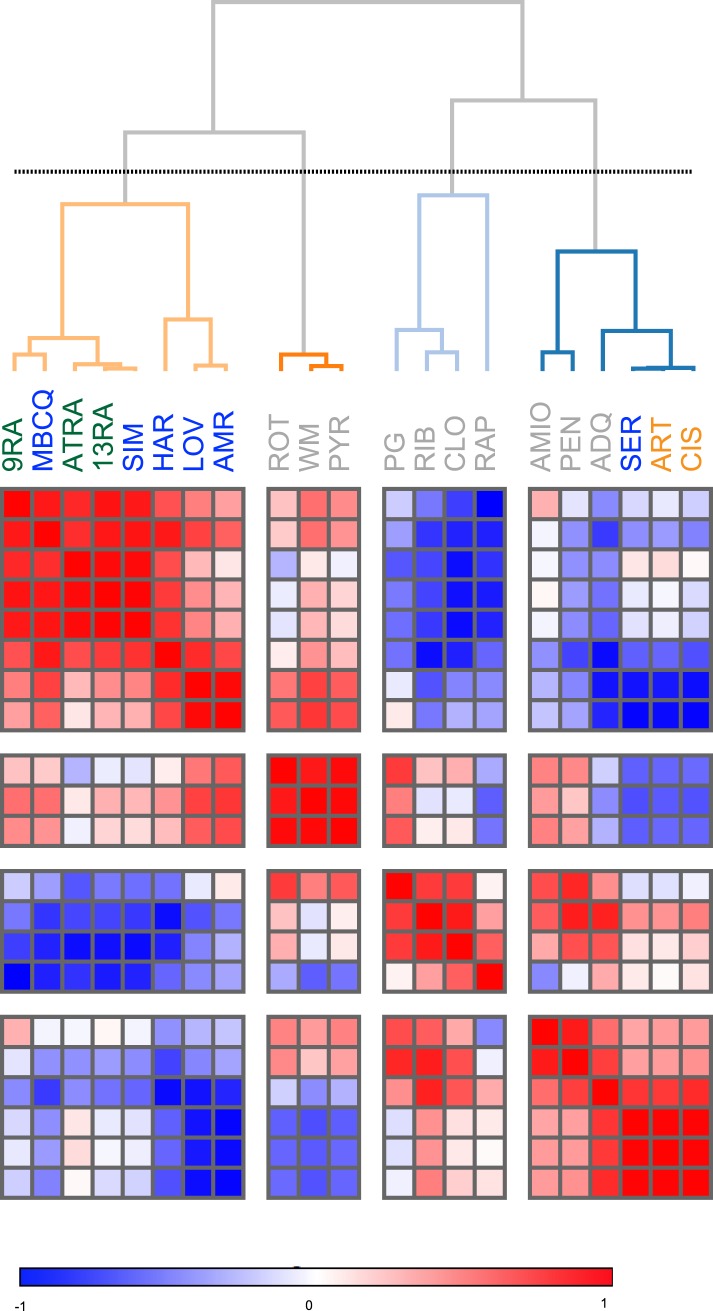
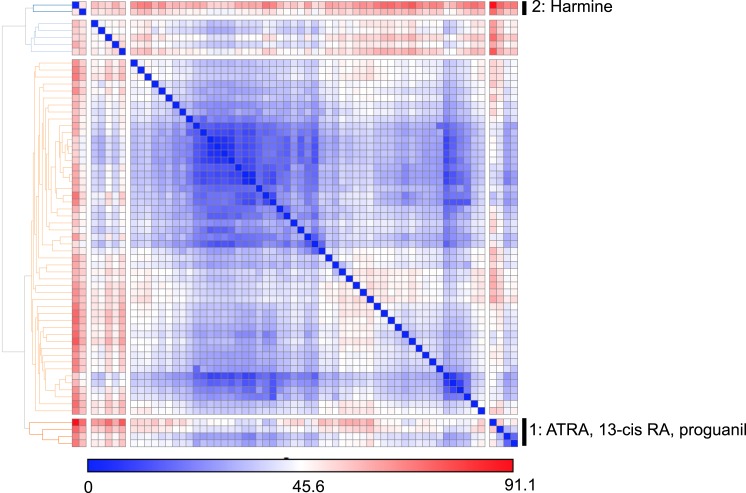
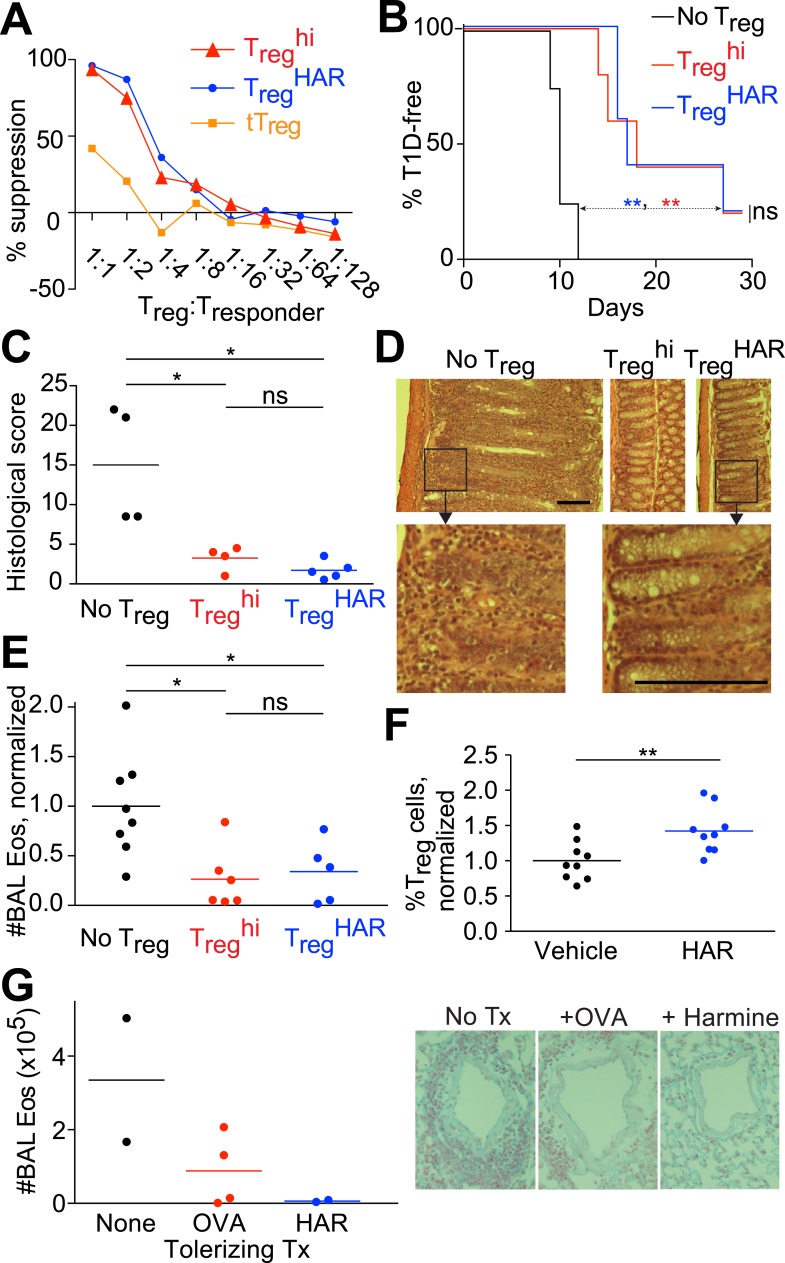
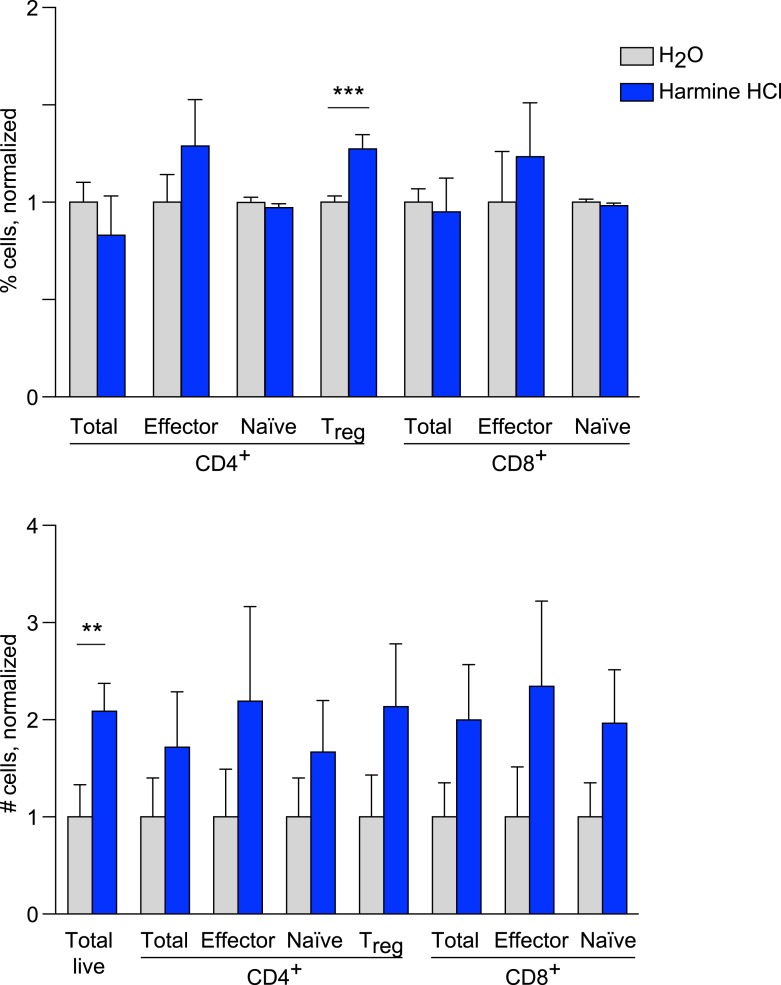
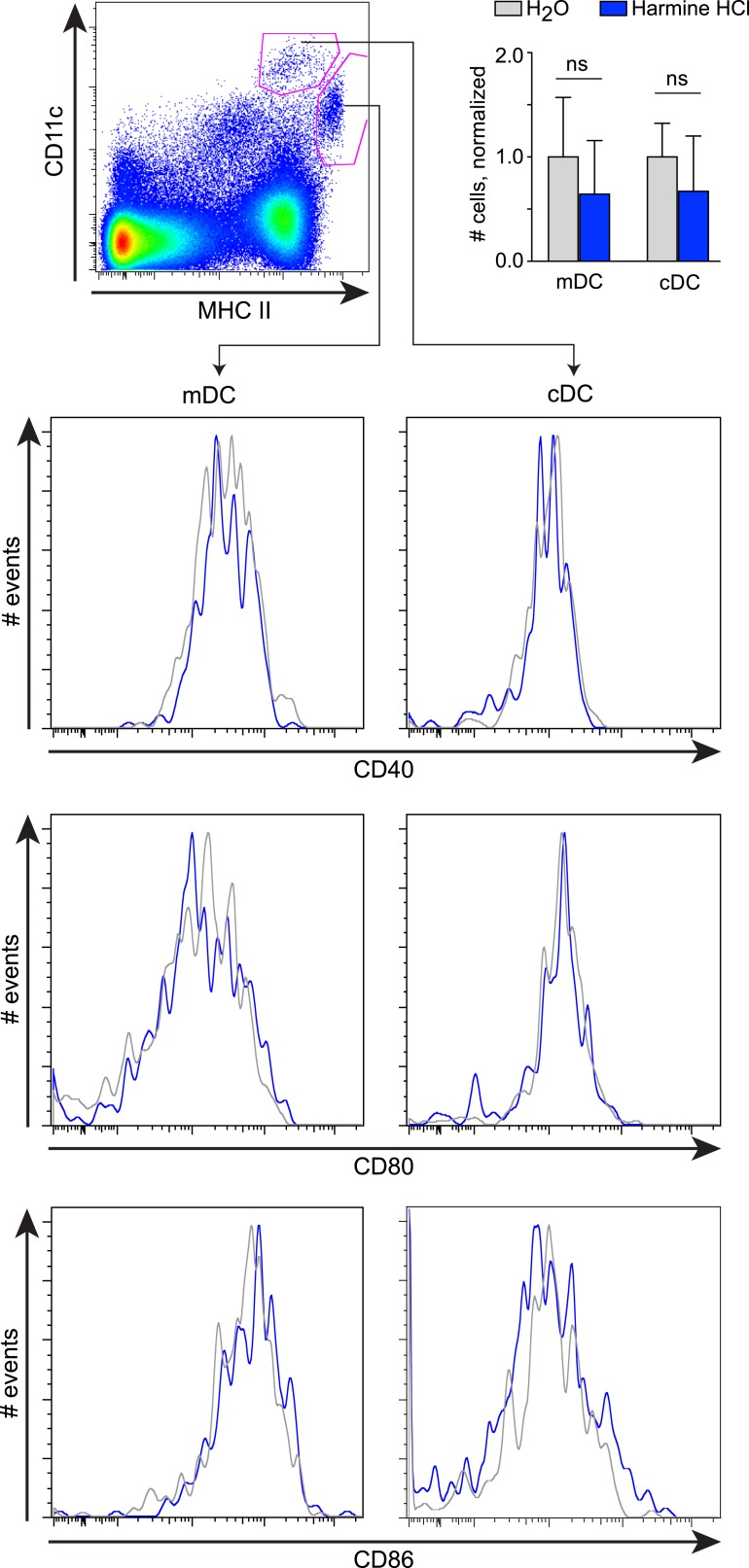
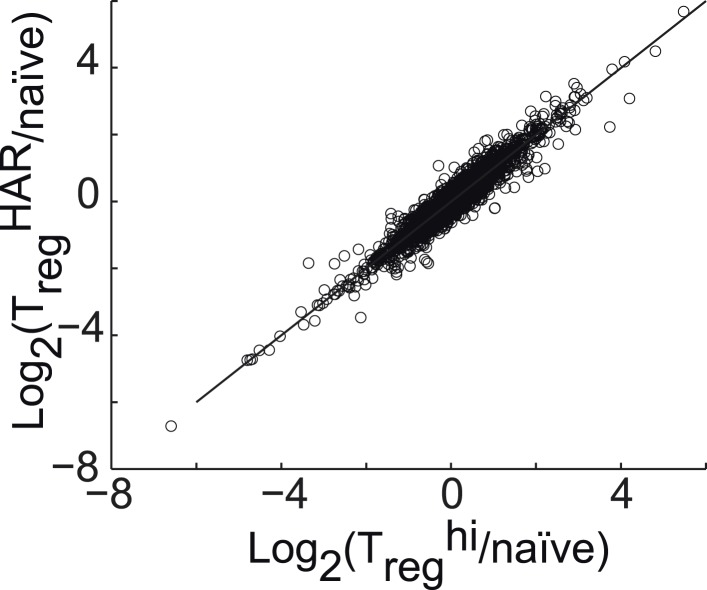
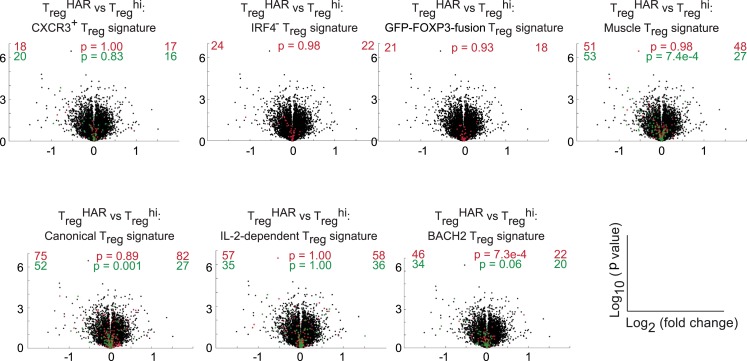
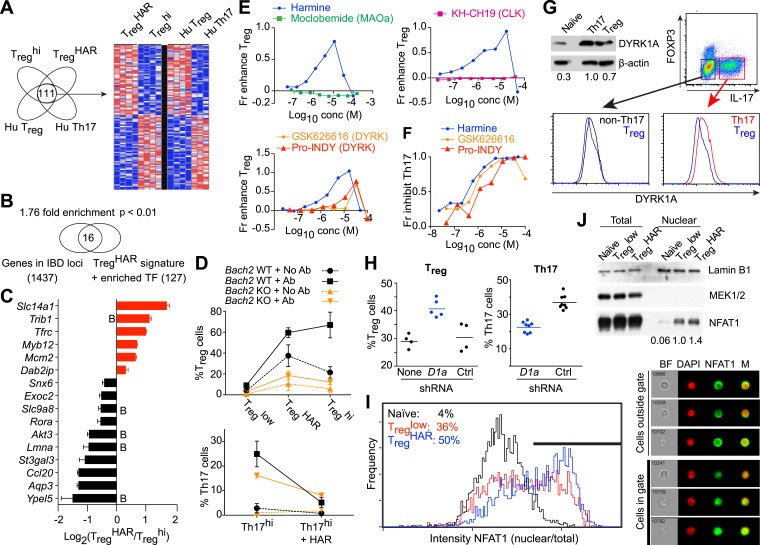
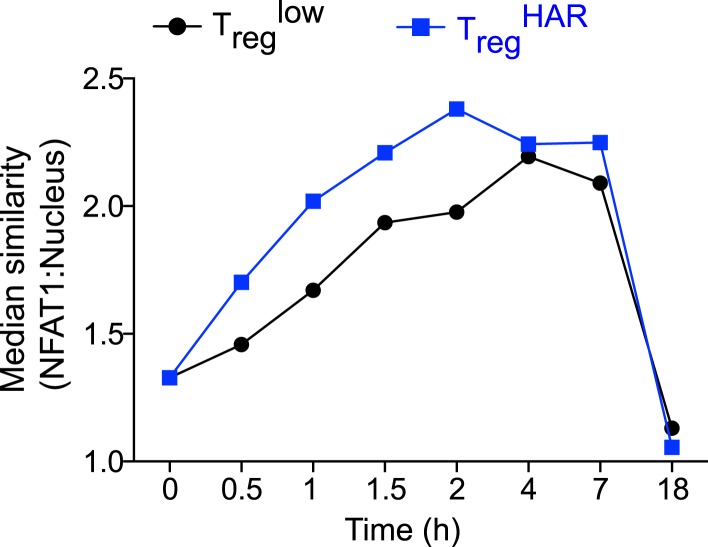
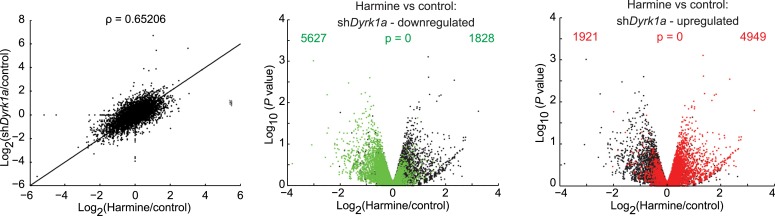
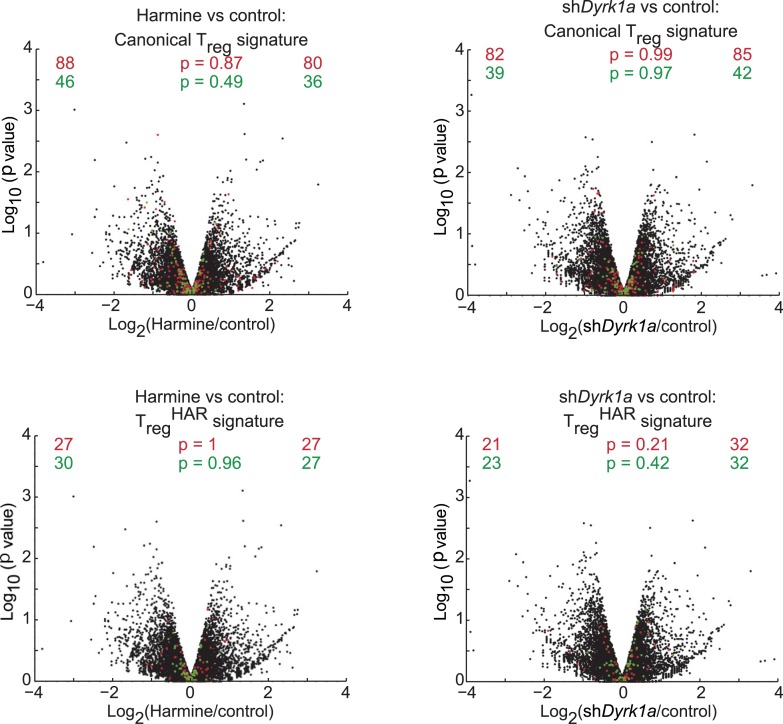
References
Publication types
MeSH terms
Substances
Grants and funding
- T32 HL066987/HL/NHLBI NIH HHS/United States
- P30 DK043351/DK/NIDDK NIH HHS/United States
- T32 CA009216-31/CA/NCI NIH HHS/United States
- K08 DK104021-01/DK/NIDDK NIH HHS/United States
- 105663/Z/14/Z/WT_/Wellcome Trust/United Kingdom
- T32 CA009216/CA/NCI NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- K08 DK104021/DK/NIDDK NIH HHS/United States
- 105663/WT_/Wellcome Trust/United Kingdom
- S10 OD012027/OD/NIH HHS/United States
- T32 HL066987-12/HL/NHLBI NIH HHS/United States
- S10 OD012027-01A1/OD/NIH HHS/United States
- U01 DK062432/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
