

Phosphorylation of TGB1 by protein kinase CK2 promotes barley stripe mosaic virus movement in monocots and dicots
- PMID: 25998907
- PMCID: PMC4507770
- DOI: 10.1093/jxb/erv237
Phosphorylation of TGB1 by protein kinase CK2 promotes barley stripe mosaic virus movement in monocots and dicots
Abstract
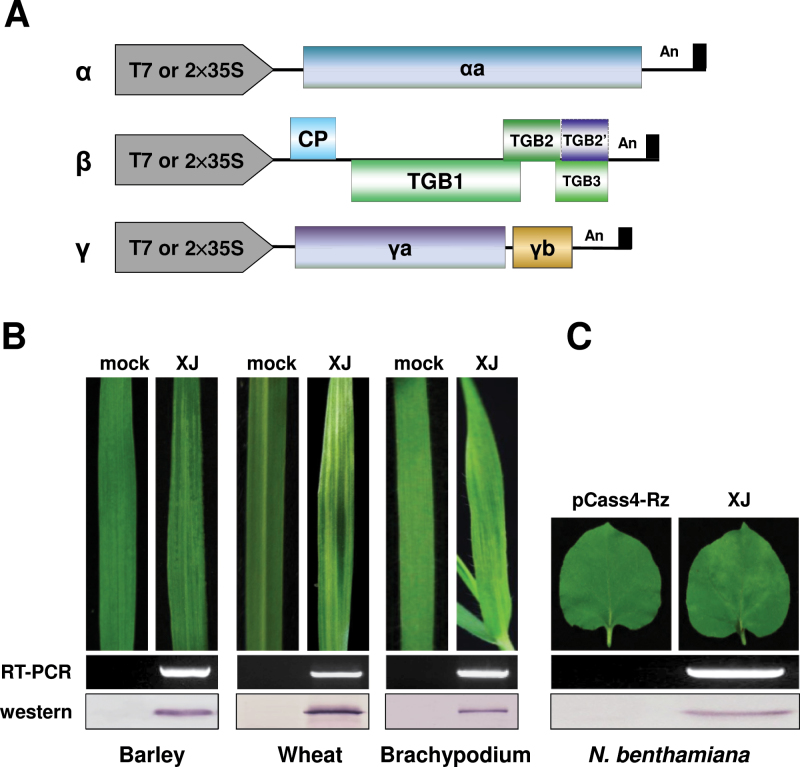
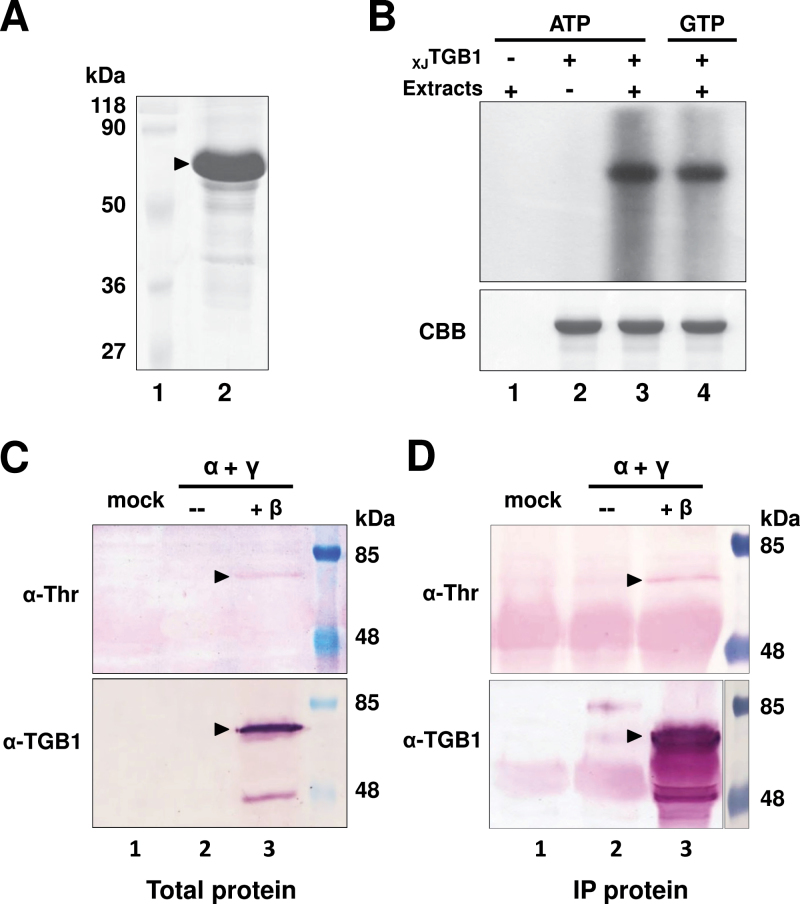
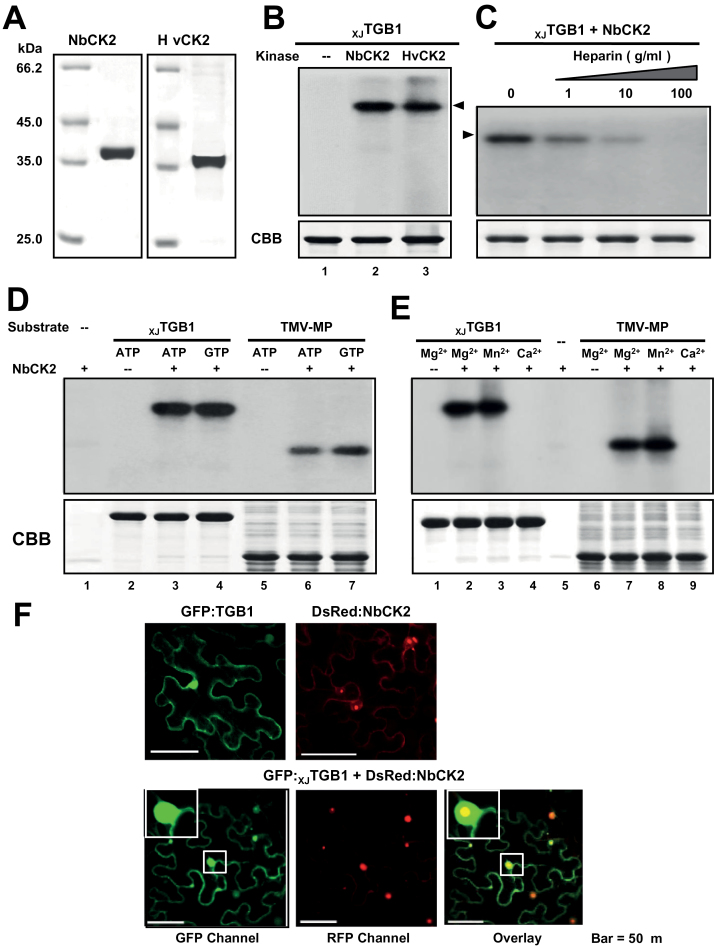
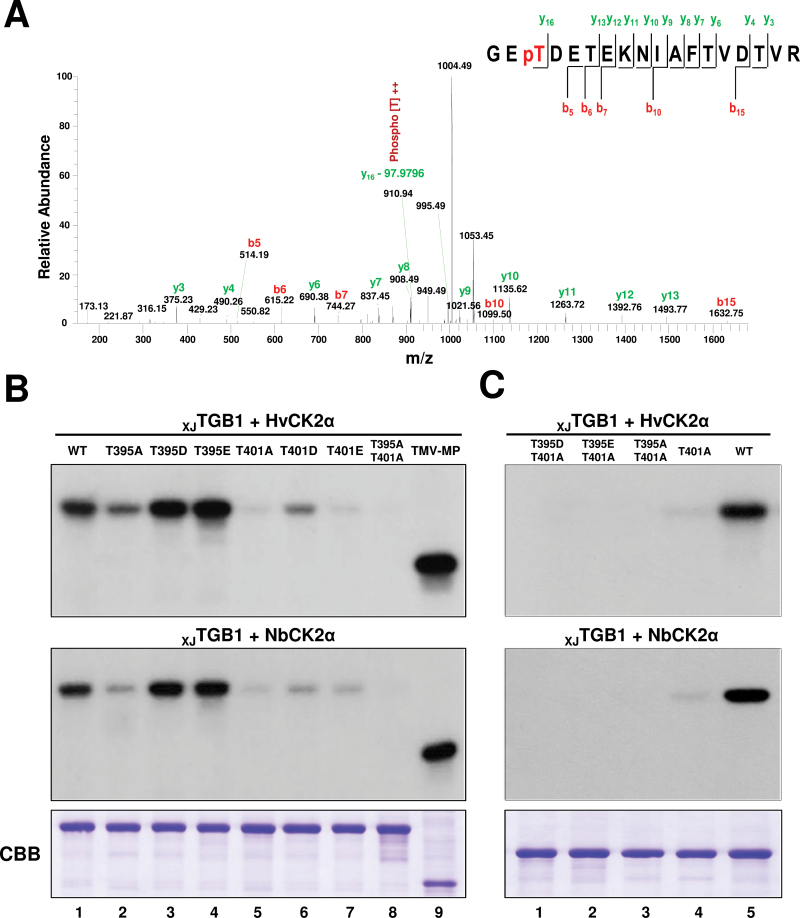
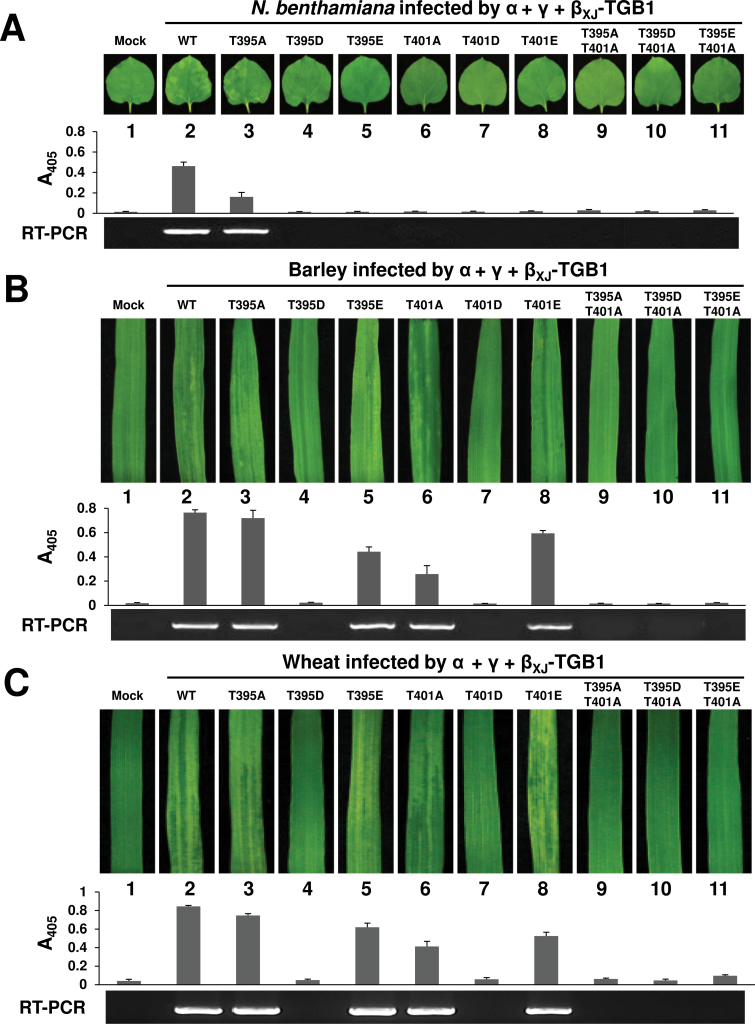
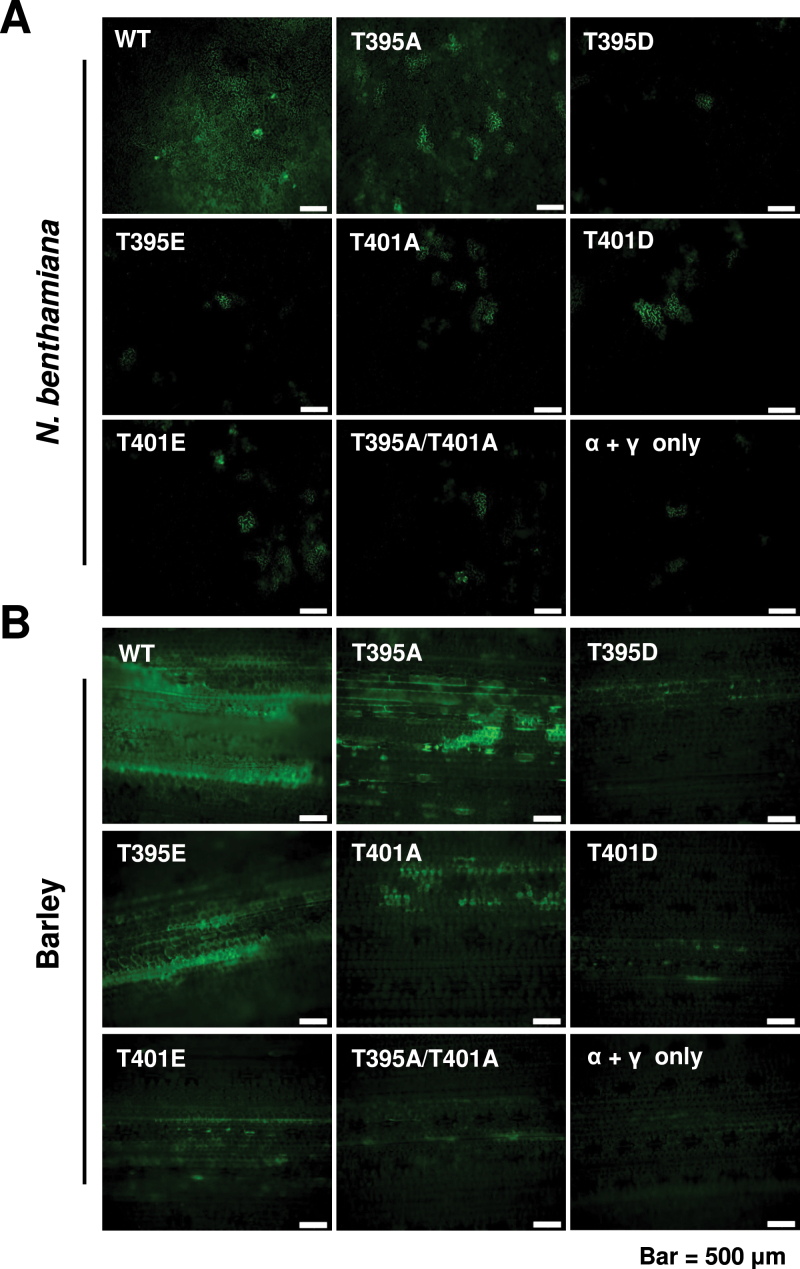
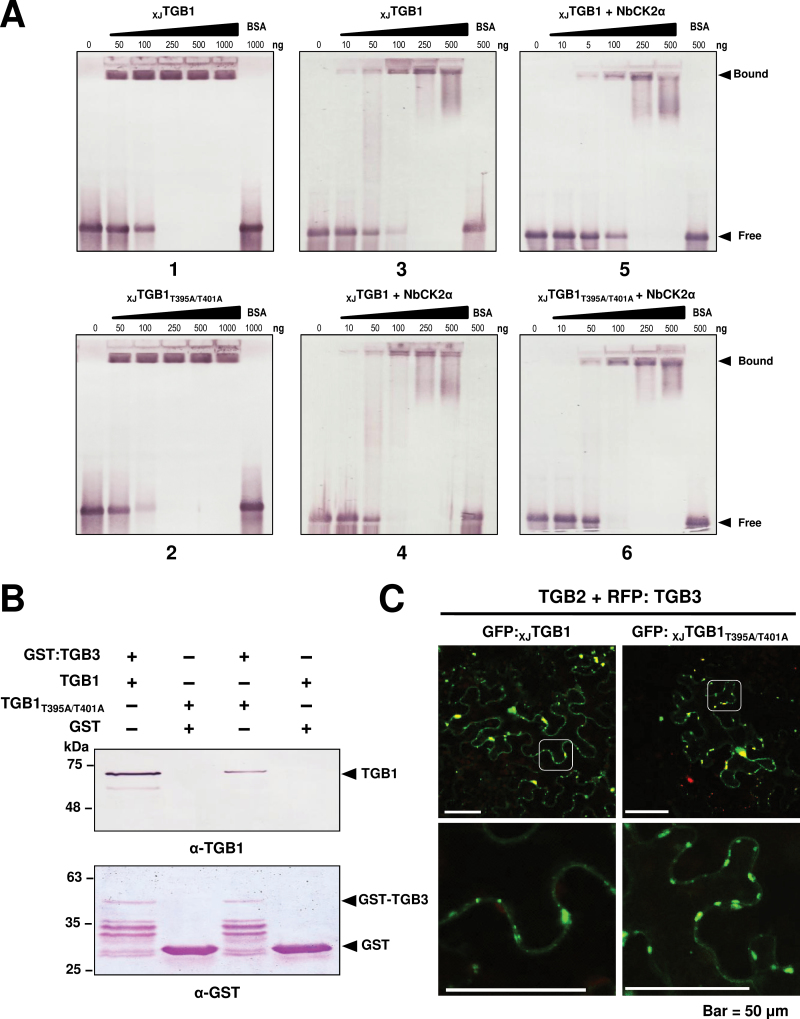
The barley stripe mosaic virus (BSMV) triple gene block 1 (TGB1) protein is required for virus cell-to-cell movement. However, little information is available about how these activities are regulated by post-translational modifications. In this study, we showed that the BSMV Xinjiang strain TGB1 (XJTGB1) is phosphorylated in vivo and in vitro by protein kinase CK2 from barley and Nicotiana benthamiana. Liquid chromatography tandem mass spectrometry analysis and in vitro phosphorylation assays demonstrated that Thr-401 is the major phosphorylation site of the XJTGB1 protein, and suggested that a Thr-395 kinase docking site supports Thr-401 phosphorylation. Substitution of Thr-395 with alanine (T395A) only moderately impaired virus cell-to-cell movement and systemic infection. In contrast, the Thr-401 alanine (T401A) virus mutant was unable to systemically infect N. benthamiana but had only minor effects in monocot hosts. Substitution of Thr-395 or Thr-401 with aspartic acid interfered with monocot and dicot cell-to-cell movement and the plants failed to develop systemic infections. However, virus derivatives with single glutamic acid substitutions at Thr-395 and Thr-401 developed nearly normal systemic infections in the monocot hosts but were unable to infect N. benthamiana systemically, and none of the double mutants was able to infect dicot and monocot hosts. The mutant XJTGB1T395A/T401A weakened in vitro interactions between XJTGB1 and XJTGB3 proteins but had little effect on XJTGB1 RNA-binding ability. Taken together, our results support a critical role of CK2 phosphorylation in the movement of BSMV in monocots and dicots, and provide new insights into the roles of phosphorylation in TGB protein functions.
Keywords: Barley stripe mosaic virus; phosphorylation; promotion; protein kinase CK2; triple gene block 1 (TGB1) protein; viral movement..
© The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Akamatsu N, Takeda A, Kishimoto M, Kaido M, Okuno T, Mise K. 2007. Phosphorylation and interaction of the movement and coat proteins of brome mosaic virus in infected barley protoplasts. Archives of Virology 152, 2087–2093. - PubMed
-
- Annamalai P, Rao ALN. 2005. Replication-independent expression of genome components and capsid protein of Brome mosaic virus in planta: a functional role for viral replicase in RNA packaging. Virology 338, 96–111. - PubMed
-
- Battistutta R, Sarno S, De Moliner E, Marin O, Issinger O-G, Zanotti G, Pinna LA. 2000. The crystal structure of the complex of Zea mays α subunit with a fragment of human β subunit provides the clue to the architecture of protein kinase CK2 holoenzyme. European Journal of Biochemistry 267, 5184–5190. - PubMed
-
- Bombarely A, Rosli HG, Vrebalov J, Moffett P, Mueller LA, Martin GB. 2012. A draft genome sequence of Nicotiana benthamiana to enhance molecular plant–microbe biology research. Molecular Plant–Microbe Interactions 25, 1523–1530. - PubMed
-
- Bond AE, Row PE, Dudley E. 2011. Post-translation modification of proteins; methodologies and applications in plant sciences. Phytochemistry 72, 975–996. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
