

doi: 10.1016/j.chembiol.2015.04.013.
Peroxide-Dependent MGL Sulfenylation Regulates 2-AG-Mediated Endocannabinoid Signaling in Brain Neurons
Affiliations
- PMID: 26000748
- PMCID: PMC4459529
- DOI: 10.1016/j.chembiol.2015.04.013
Item in Clipboard
Peroxide-Dependent MGL Sulfenylation Regulates 2-AG-Mediated Endocannabinoid Signaling in Brain Neurons
Chem Biol.
.
Abstract
The second messenger hydrogen peroxide transduces changes in the cellular redox state by reversibly oxidizing protein cysteine residues to sulfenic acid. This signaling event regulates many cellular processes but has never been shown to occur in the brain. Here, we report that hydrogen peroxide heightens endocannabinoid signaling in brain neurons through sulfenylation of cysteines C201 and C208 in monoacylglycerol lipase (MGL), a serine hydrolase that deactivates the endocannabinoid 2-arachidonoyl-sn-glycerol (2-AG) in nerve terminals. The results suggest that MGL sulfenylation may provide a presynaptic control point for 2-AG-mediated endocannabinoid signaling.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
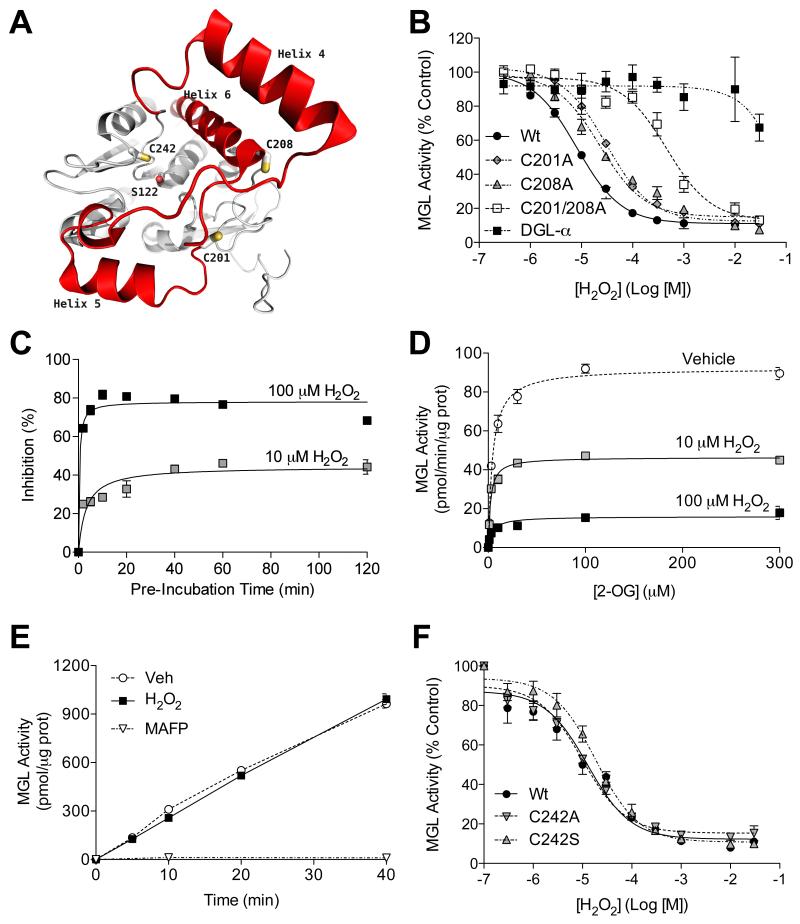
(A) Structure of human MGL showing cysteine residues C201 and C208 (on the cap domain, red) and C242 (close to catalytic nucleophile S122). (B) Effects of H2O2 on wild-type MGL (filled circles), MGL mutants C201A (filled diamonds), C208A (filled triangles) or C201/208A (empty squares) and DGL-α (filled squares). (C) Time-course of MGL inhibition by H2O2 (10 μM: shaded squares; 100 μM: filled squares). (D) Kinetics of MGL inhibition by H2O2 (vehicle: open circles). (E) Rapid dilution of MGL incubated with vehicle (circles), H2O2 (filled squares; 10 μM final), or irreversible inhibitor methylarachidonylfluorophosphonate (MAFP, inverted triangles, 23 nM final). (F) Effects of H2O2 on wild-type MGL (filled circles) and MGL C242 mutants C242A (inverted triangles) and C242S (triangles).
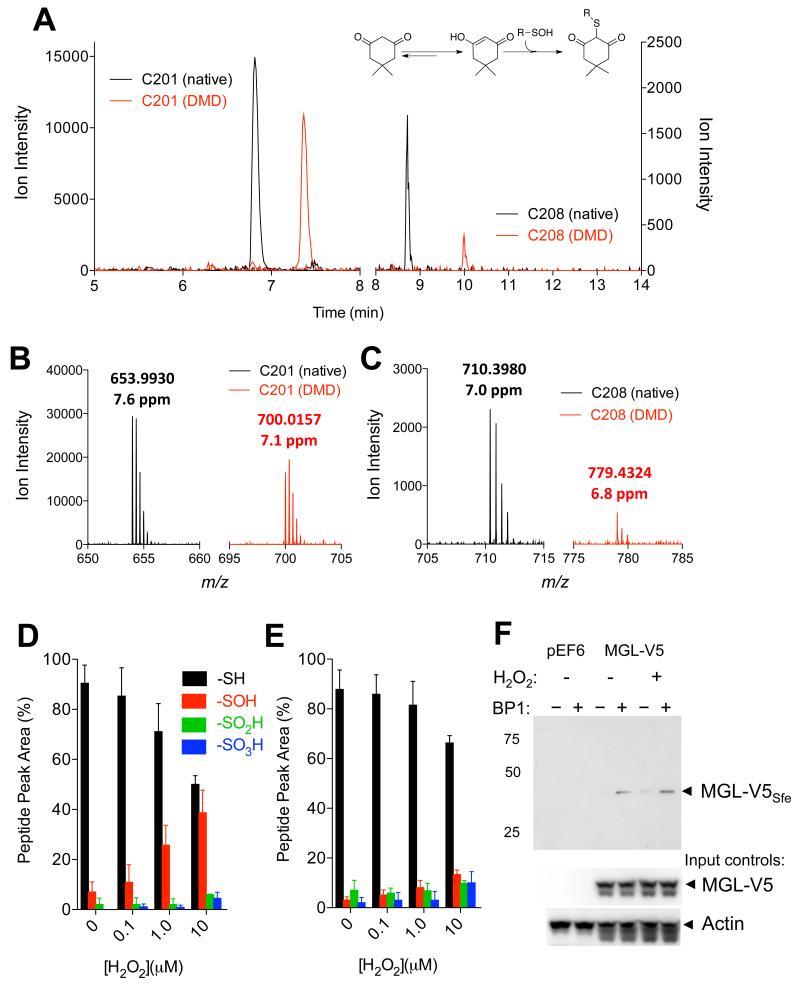
(A) Extracted-ion currents of native (black) and dimedone (DMD)-containing (red) peptides obtained by digestion of rat MGL exposed to H2O2 (10 μM, 1h). Inset: reaction of DMD with sulfenylated cysteine. (B,C) Mass spectra regions showing that DMD binds to C201 (B) and C208 (C). The observed m/z values and the corresponding mass errors are listed above the peaks. (D,E) Relative abundance of C201 and C208 present as native (-SH), sulfenic (-SOH, dimedone adduct), sulfinic (-SO2H) and sulfonic acid (-SO3H) at varying H2O2 concentrations. (F) Detection of sulfenylated MGL (MGL-V5Sfe) in Neuro-2a cells after H2O2 treatment (100 μM, 1h). Sulfenylated MGL was trapped using the chemoselective probe biotin-1,3-cyclopentanedione (BP1). Equal sample loading to the affinity precipitation was confirmed by assessing levels of MGL (using an anti-V5 antibody) and actin (input controls). 75, 50, 25: molecular weight size markers (kDa).
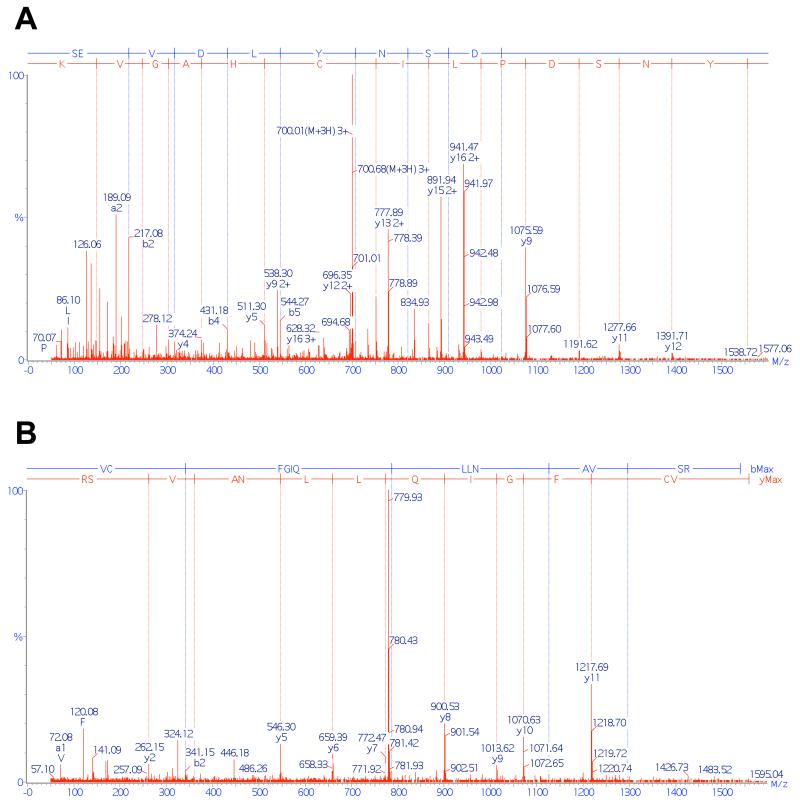
(A) Tandem mass spectra of peptide SEVDLYNSDPLICHAGVK, showing that sulfenylation (detected as DMD adduct) occurred on C201. (B) Tandem mass spectra of peptide VCFGIQLLNAVSR, showing that sulfenylation (detected as DMD adduct) occurred on C208.
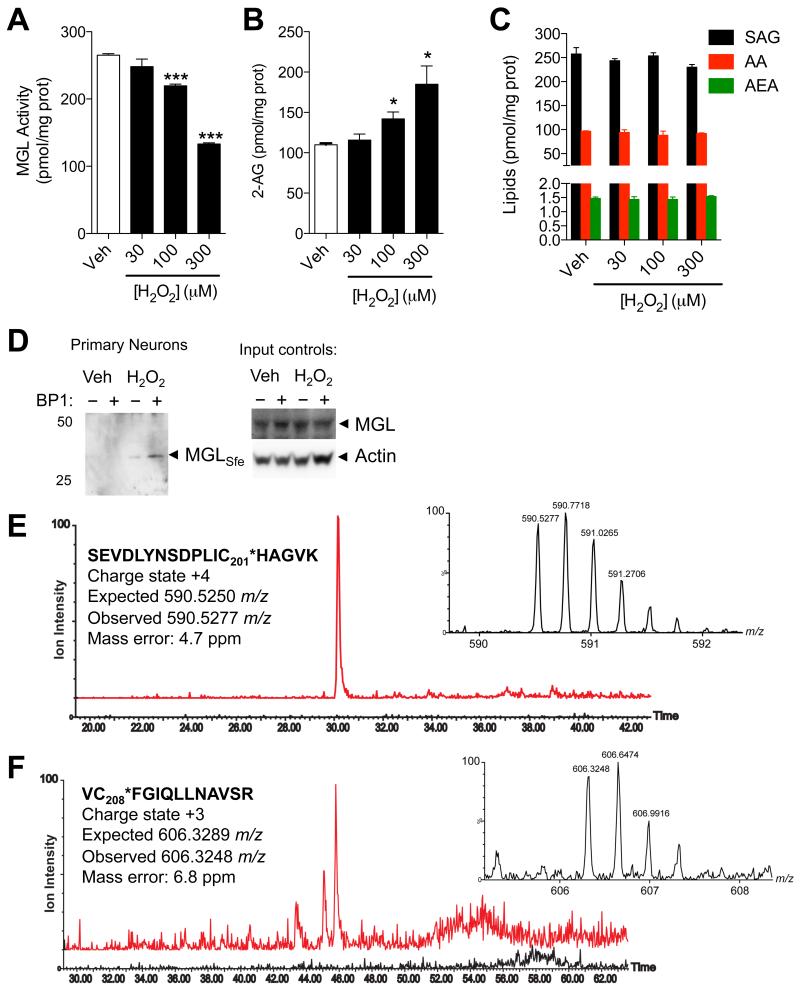
(A-D) Effects of H2O2 (filled bars) or vehicle (open bars) on (A) MGL activity, (B) 2-AG levels, and (C) 1-stearoyl,2-arachidonoylglycerol (SAG), arachidonic acid (AA), and anandamide (AEA) in rat cortical neurons in primary cultures. (D) Detection of sulfenylated MGL (MGLSfe) in primary cortical neurons in cultures after H2O2 treatment (300 μM, 1h). Sulfenylated MGL was trapped using the chemoselective probe BP1. Equal sample loading to the affinity precipitation was confirmed by measuring MGL (anti-MGL antibody) and actin (input controls). 50, 25: molecular weight size markers (kDa). (E) Ion current of the BP1-adduct of MGL peptide bearing C201 (590.53 m/z, z=4) extracted from the control incubation (black trace) and incubation of neurons with H2O2 (red trace). (F) Ion current of the BP1-adduct of MGL peptide bearing C208 (606.30 m/z, z=3) extracted from the control incubation (black trace) and incubation of neurons with H2O2 (red trace). In both E and F, the high-resolution mass spectra reported in the inset matches the expected charge state and m/z value. ***P<0.001 and *P<0.05 compared to vehicle, two-tailed Student’s t test.
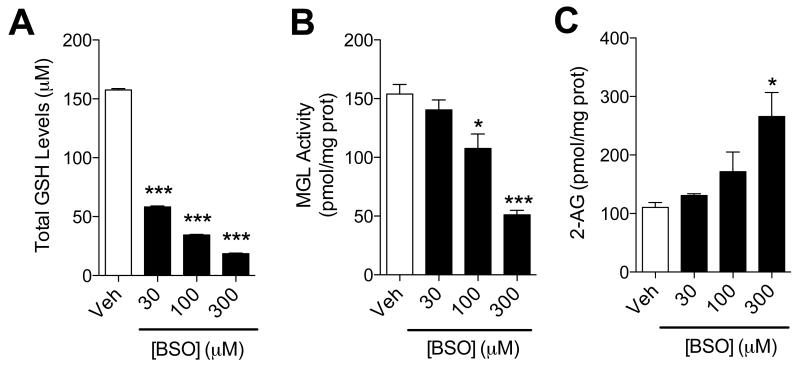
(A-C) Effects of BSO (filled bars) or vehicle (open bars) on (A) GSH levels, (B) MGL activity and (C) 2-AG levels in neuronal cultures. ***P<0.001 and *P<0.05 compared to vehicle, two-tailed Student’s t test.
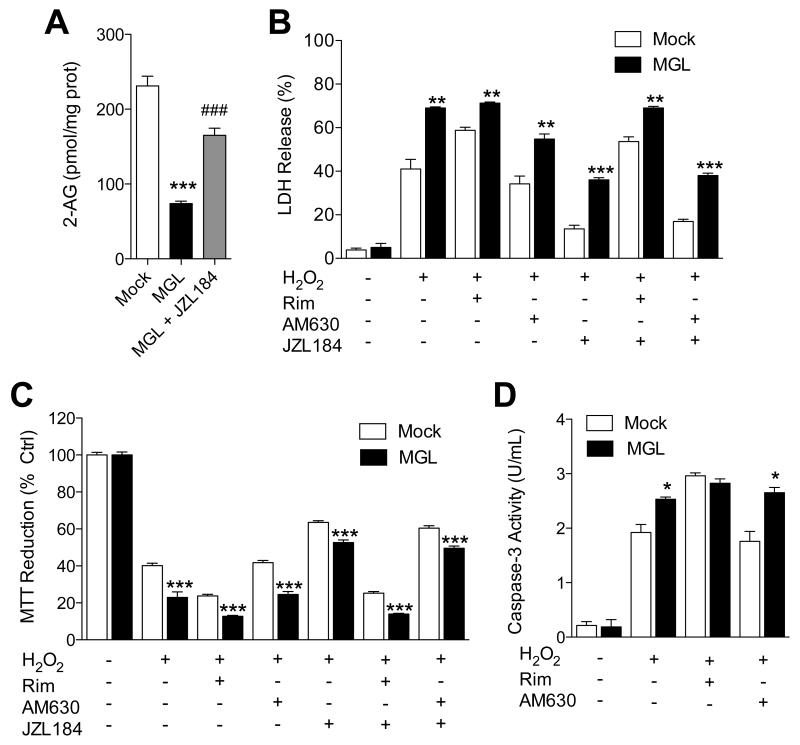
(A) 2-AG levels in Neuro-2a cells overexpressing MGL. (B-D) Effects of H2O2, alone or combined with CB1 antagonist rimonabant, CB2 antagonist AM630 or MGL inhibitor JZL184 (each at 1 μM), on wild-type (open bars) or MGL-overexpressing Neuro-2a cells: (B) LDH release, (C) MTT reduction, and (D) caspase-3 activity. ***P<0.001, **P<0.05 and * P<0.05 compared to Mock, and ###P<0.001 compared to MGL, two-tailed Student’s t test.
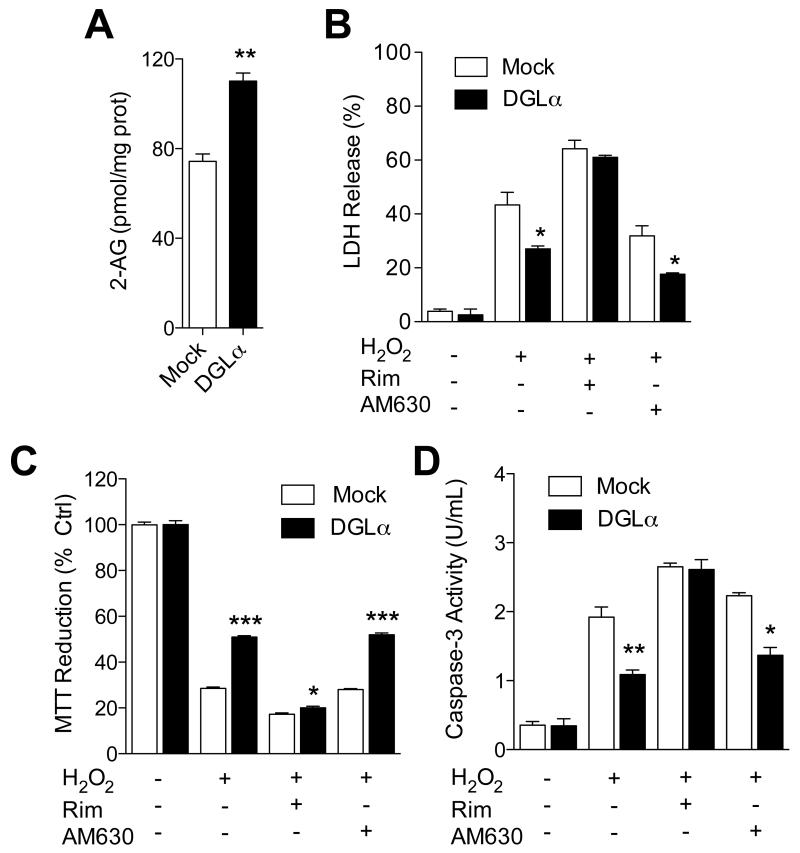
(A) 2-AG levels in Neuro-2a cells overexpressing DGL-α. (B-D) Effects of H2O2, alone or combined with rimonabant, AM630 or JZL184, on wild-type or DGL-α–overexpressing Neuro-2a cells: (B) LDH release, (C) MTT reduction, and (D) caspase-3 activity. ***P<0.001, **P<0.05 and * P<0.05 compared to Mock, two-tailed Student’s t test.
References
-
- Bénard G, Massa F, Puente N, Lourenço J, Bellocchio L, Soria-Gómez E, Matias I, Delamarre A, Metna-Laurent M, Cannich A, Hebert-Chatelain E, Mulle C, Ortega-Gutiérrez S, Martín-Fontecha M, Klugmann M, Guggenhuber S, Lutz B, Gertsch J, Chaouloff F, López-Rodríguez ML, Grandes P, Rossignol R, Marsicano G. Mitochondrial CB1 receptors regulate neuronal energy metabolism. Nat Neurosci. 2012;15:558–564. - PubMed
-
- Bertrand T, Augé F, Houtmann J, Rak A, Vallée F, Mikol V, Berne PF, Michot N, Cheuret D, Hoornaert C, Mathieu M. Structural basis for human monoglyceride lipase inhibition. J. Mol. Biol. 2010;396:663–673. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
