

Complement Component 5 Mediates Development of Fibrosis, via Activation of Stellate Cells, in 2 Mouse Models of Chronic Pancreatitis
- PMID: 26001927
- PMCID: PMC4560830
- DOI: 10.1053/j.gastro.2015.05.012
Complement Component 5 Mediates Development of Fibrosis, via Activation of Stellate Cells, in 2 Mouse Models of Chronic Pancreatitis
Abstract
Background & aims: Little is known about the pathogenic mechanisms of chronic pancreatitis. We investigated the roles of complement component 5 (C5) in pancreatic fibrogenesis in mice and patients.
Methods: Chronic pancreatitis was induced by ligation of the midpancreatic duct, followed by a single supramaximal intraperitoneal injection of cerulein, in C57Bl6 (control) and C5-deficient mice. Some mice were given injections of 2 different antagonists of the receptor for C5a over 21 days. In a separate model, mice were given injections of cerulein for 10 weeks to induce chronic pancreatitis. Direct effects of C5 were studied in cultured primary cells. We performed genotype analysis for the single-nucleotide polymorphisms rs 17611 and rs 2300929 in C5 in patients with pancreatitis and healthy individuals (controls). Blood cells from 976 subjects were analyzed by transcriptional profiling.
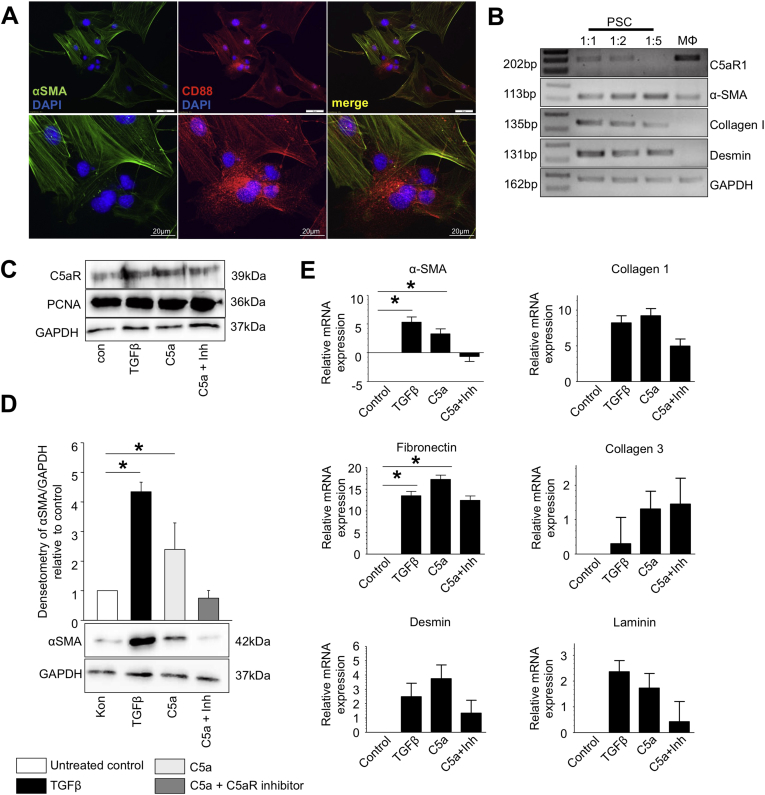
Results: During the initial phase of pancreatitis, levels of pancreatic damage were similar between C5-deficient and control mice. During later stages of pancreatitis, C5-deficient mice and mice given injections of C5a-receptor antagonists developed significantly less pancreatic fibrosis than control mice. Primary pancreatic stellate cells were activated in vitro by C5a. There were no differences in the rs 2300929 SNP between subjects with or without pancreatitis, but the minor allele rs17611 was associated with a significant increase in levels of C5 in whole blood.
Conclusions: In mice, loss of C5 or injection of a C5a-receptor antagonist significantly reduced the level of fibrosis of chronic pancreatitis, but this was not a consequence of milder disease in early stages of pancreatitis. C5 might be a therapeutic target for chronic pancreatitis.
Keywords: Complement System; Pancreatic Stellate Cells; Transcriptome Analysis; αSMA.
Copyright © 2015 AGA Institute. Published by Elsevier Inc. All rights reserved.
Figures














Similar articles
-
Retinoic Acid Ameliorates Pancreatic Fibrosis and Inhibits the Activation of Pancreatic Stellate Cells in Mice with Experimental Chronic Pancreatitis via Suppressing the Wnt/β-Catenin Signaling Pathway.PLoS One. 2015 Nov 10;10(11):e0141462. doi: 10.1371/journal.pone.0141462. eCollection 2015. PLoS One. 2015. PMID: 26556479 Free PMC article.
-
Characterization of Mouse Models of Early Pancreatic Lesions Induced by Alcohol and Chronic Pancreatitis.Pancreas. 2015 Aug;44(6):882-7. doi: 10.1097/MPA.0000000000000380. Pancreas. 2015. PMID: 26166469 Free PMC article.
-
Aryl Hydrocarbon Receptor Ligands in Cigarette Smoke Induce Production of Interleukin-22 to Promote Pancreatic Fibrosis in Models of Chronic Pancreatitis.Gastroenterology. 2016 Dec;151(6):1206-1217. doi: 10.1053/j.gastro.2016.09.064. Epub 2016 Oct 18. Gastroenterology. 2016. PMID: 27769811 Free PMC article.
-
A New Insight into Chronic Pancreatitis.Tohoku J Exp Med. 2019 Aug;248(4):225-238. doi: 10.1620/tjem.248.225. Tohoku J Exp Med. 2019. PMID: 31378749 Review.
-
Pancreatic Stellate Cells and the Targeted Therapeutic Strategies in Chronic Pancreatitis.Molecules. 2023 Jul 22;28(14):5586. doi: 10.3390/molecules28145586. Molecules. 2023. PMID: 37513458 Free PMC article. Review.
Cited by
-
Therapeutic Intervention with Anti-Complement Component 5 Antibody Does Not Reduce NASH but Does Attenuate Atherosclerosis and MIF Concentrations in Ldlr-/-.Leiden Mice.Int J Mol Sci. 2022 Sep 14;23(18):10736. doi: 10.3390/ijms231810736. Int J Mol Sci. 2022. PMID: 36142647 Free PMC article.
-
Animal models of gastrointestinal and liver diseases. Animal models of acute and chronic pancreatitis.Am J Physiol Gastrointest Liver Physiol. 2016 Sep 1;311(3):G343-55. doi: 10.1152/ajpgi.00372.2015. Epub 2016 Jul 14. Am J Physiol Gastrointest Liver Physiol. 2016. PMID: 27418683 Free PMC article. Review.
-
5-FU-miR-15a Inhibits Activation of Pancreatic Stellate Cells by Reducing YAP1 and BCL-2 Levels In Vitro.Int J Mol Sci. 2023 Feb 16;24(4):3954. doi: 10.3390/ijms24043954. Int J Mol Sci. 2023. PMID: 36835366 Free PMC article.
-
Chronic Pancreatitis and the Development of Pancreatic Cancer.Endocr Metab Immune Disord Drug Targets. 2020;20(8):1182-1210. doi: 10.2174/1871530320666200423095700. Endocr Metab Immune Disord Drug Targets. 2020. PMID: 32324526 Free PMC article. Review.
-
Cathepsin B Activity Initiates Apoptosis via Digestive Protease Activation in Pancreatic Acinar Cells and Experimental Pancreatitis.J Biol Chem. 2016 Jul 8;291(28):14717-31. doi: 10.1074/jbc.M116.718999. Epub 2016 May 12. J Biol Chem. 2016. PMID: 27226576 Free PMC article.
References
-
- Gress T.M., Muller-Pillasch F., Lerch M.M., et al. Balance of expression of genes coding for extracellular matrix proteins and extracellular matrix degrading proteases in chronic pancreatitis. Z Gastroenterol. 1994;32:221–225. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
