

Constructing the suprachiasmatic nucleus: a watchmaker's perspective on the central clockworks
- PMID: 26005407
- PMCID: PMC4424844
- DOI: 10.3389/fnsys.2015.00074
Constructing the suprachiasmatic nucleus: a watchmaker's perspective on the central clockworks
Abstract
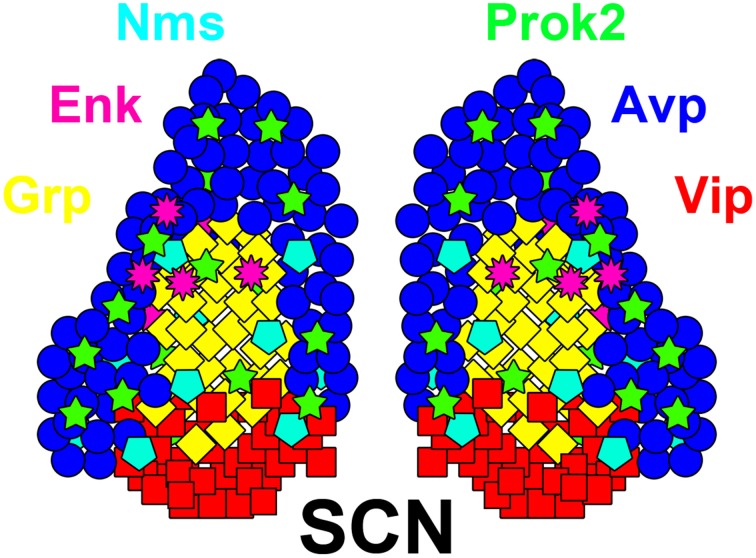
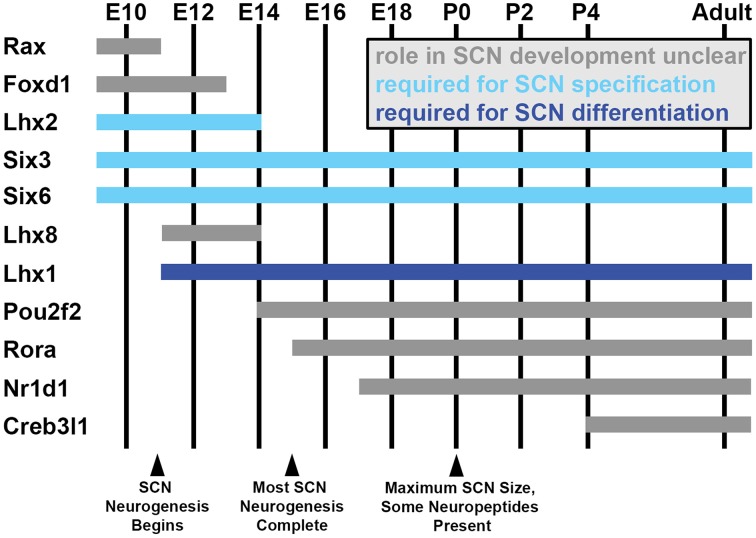
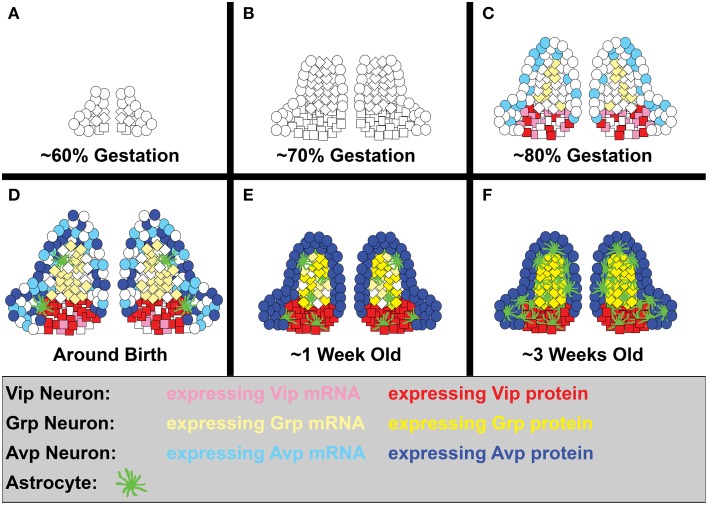
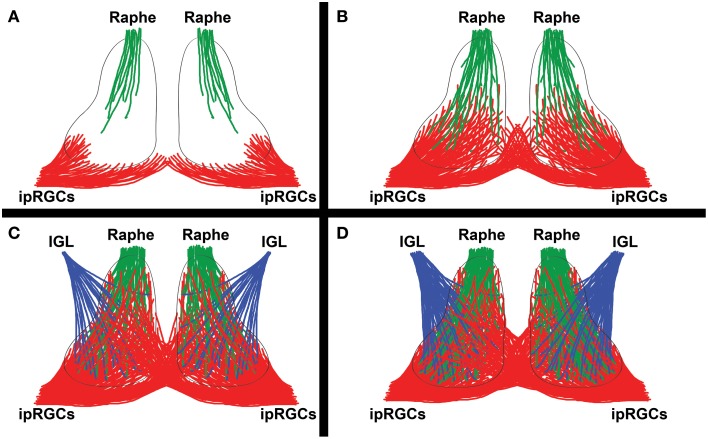
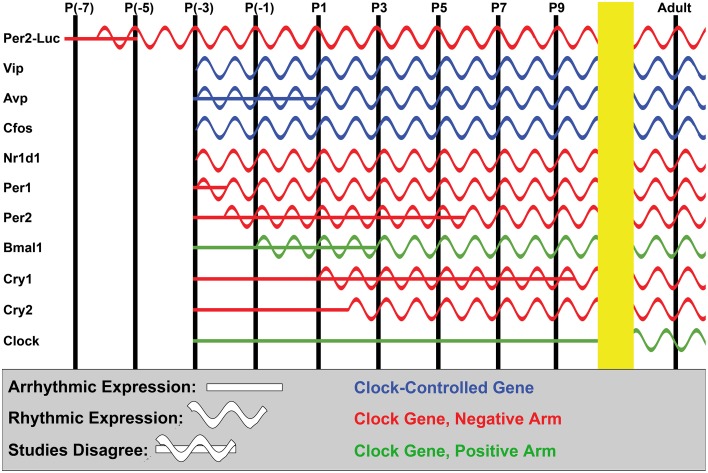
The circadian system constrains an organism's palette of behaviors to portions of the solar day appropriate to its ecological niche. The central light-entrained clock in the suprachiasmatic nucleus (SCN) of the mammalian circadian system has evolved a complex network of interdependent signaling mechanisms linking multiple distinct oscillators to serve this crucial function. However, studies of the mechanisms controlling SCN development have greatly lagged behind our understanding of its physiological functions. We review advances in the understanding of adult SCN function, what has been described about SCN development to date, and the potential of both current and future studies of SCN development to yield important insights into master clock function, dysfunction, and evolution.
Keywords: axon pathfinding; cell fate; chronotype; circadian; clock genes; developmental neuroscience; suprachiasmatic; transcription factors.
Figures
References
-
- Abrahamson E. E., Moore R. Y. (2001). Suprachiasmatic nucleus in the mouse: retinal innervation, intrinsic organization and efferent projections. Brain Res. 916, 172–191. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources