

Calcineurin orchestrates dimorphic transitions, antifungal drug responses and host-pathogen interactions of the pathogenic mucoralean fungus Mucor circinelloides
- PMID: 26010100
- PMCID: PMC4633022
- DOI: 10.1111/mmi.13071
Calcineurin orchestrates dimorphic transitions, antifungal drug responses and host-pathogen interactions of the pathogenic mucoralean fungus Mucor circinelloides
Abstract
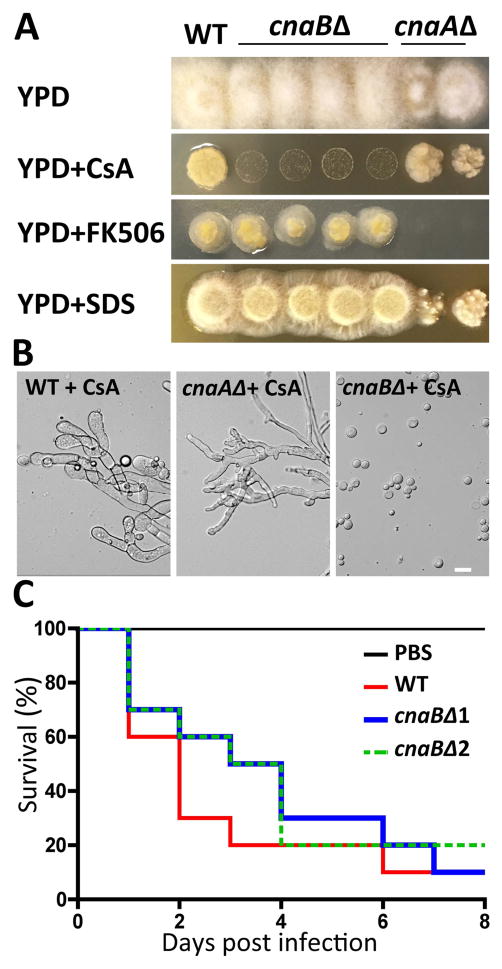
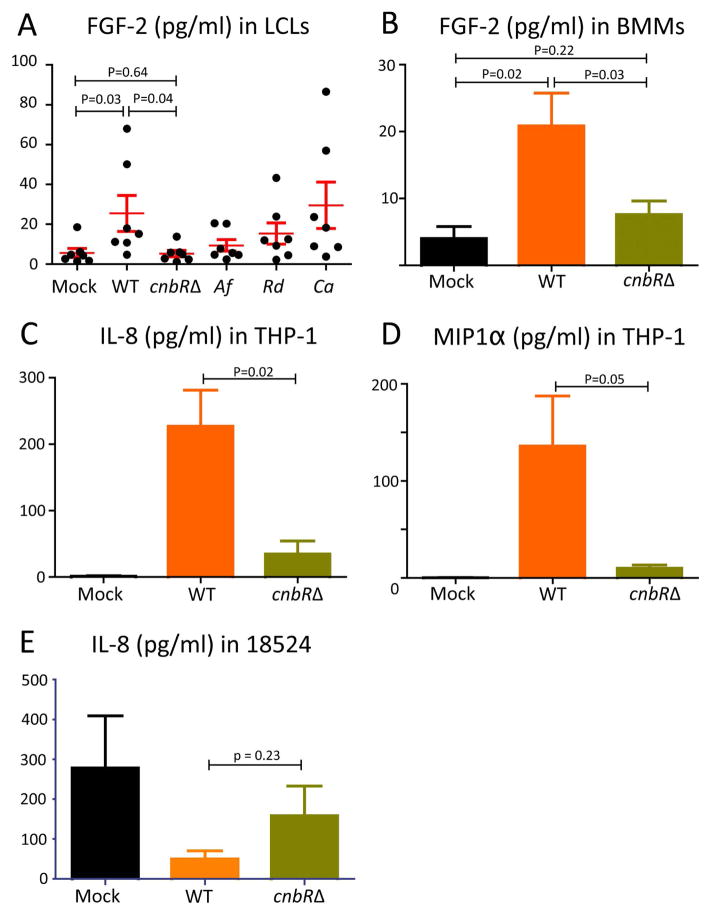
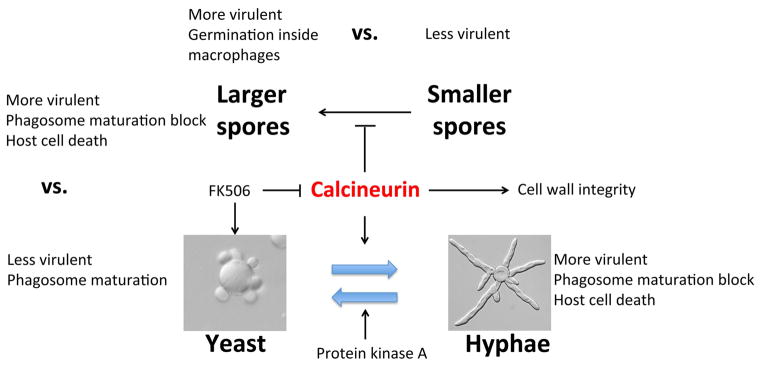
Calcineurin plays essential roles in virulence and growth of pathogenic fungi and is a target of the natural products FK506 and Cyclosporine A. In the pathogenic mucoralean fungus Mucor circinelloides, calcineurin mutation or inhibition confers a yeast-locked phenotype indicating that calcineurin governs the dimorphic transition. Genetic analysis in this study reveals that two calcineurin A catalytic subunits (out of three) are functionally diverged. Homology modeling illustrates modes of resistance resulting from amino substitutions in the interface between each calcineurin subunit and the inhibitory drugs. In addition, we show how the dimorphic transition orchestrated by calcineurin programs different outcomes during host-pathogen interactions. For example, when macrophages phagocytose Mucor yeast, subsequent phagosomal maturation occurs, indicating host cells respond appropriately to control the pathogen. On the other hand, upon phagocytosis of spores, macrophages fail to form mature phagosomes. Cytokine production from immune cells differs following exposure to yeast versus spores (which germinate into hyphae). Thus, the morphogenic transition can be targeted as an efficient treatment option against Mucor infection. In addition, genetic analysis (including gene disruption and mutational studies) further strengthens the understanding of calcineurin and provides a foundation to develop antifungal agents targeting calcineurin to deploy against Mucor and other pathogenic fungi.
© 2015 John Wiley & Sons Ltd.
Figures






Similar articles
-
Calcineurin plays key roles in the dimorphic transition and virulence of the human pathogenic zygomycete Mucor circinelloides.PLoS Pathog. 2013;9(9):e1003625. doi: 10.1371/journal.ppat.1003625. Epub 2013 Sep 5. PLoS Pathog. 2013. PMID: 24039585 Free PMC article.
-
A Novel Resistance Pathway for Calcineurin Inhibitors in the Human-Pathogenic Mucorales Mucor circinelloides.mBio. 2020 Jan 28;11(1):e02949-19. doi: 10.1128/mBio.02949-19. mBio. 2020. PMID: 31992620 Free PMC article.
-
Leveraging Fungal and Human Calcineurin-Inhibitor Structures, Biophysical Data, and Dynamics To Design Selective and Nonimmunosuppressive FK506 Analogs.mBio. 2021 Dec 21;12(6):e0300021. doi: 10.1128/mBio.03000-21. Epub 2021 Nov 23. mBio. 2021. PMID: 34809463 Free PMC article.
-
Calcineurin in fungal virulence and drug resistance: Prospects for harnessing targeted inhibition of calcineurin for an antifungal therapeutic approach.Virulence. 2017 Feb 17;8(2):186-197. doi: 10.1080/21505594.2016.1201250. Epub 2016 Jun 20. Virulence. 2017. PMID: 27325145 Free PMC article. Review.
-
Good fungi gone bad: the corruption of calcineurin.Bioessays. 2002 Oct;24(10):894-903. doi: 10.1002/bies.10157. Bioessays. 2002. PMID: 12325122 Review.
Cited by
-
Combinatorial strategies for combating invasive fungal infections.Virulence. 2017 Feb 17;8(2):169-185. doi: 10.1080/21505594.2016.1196300. Epub 2016 Jun 7. Virulence. 2017. PMID: 27268286 Free PMC article. Review.
-
H3K4 methylation regulates development, DNA repair, and virulence in Mucorales.IMA Fungus. 2024 Mar 14;15(1):6. doi: 10.1186/s43008-023-00136-3. IMA Fungus. 2024. PMID: 38481304 Free PMC article.
-
FKBP12-Dependent Inhibition of Calcineurin Mediates Immunosuppressive Antifungal Drug Action in Malassezia.mBio. 2017 Oct 24;8(5):e01752-17. doi: 10.1128/mBio.01752-17. mBio. 2017. PMID: 29066552 Free PMC article.
-
Construction of a Recyclable Genetic Marker and Serial Gene Deletions in the Human Pathogenic Mucorales Mucor circinelloides.G3 (Bethesda). 2017 Jul 5;7(7):2047-2054. doi: 10.1534/g3.117.041095. G3 (Bethesda). 2017. PMID: 28476909 Free PMC article.
-
Recent Advances and Future Directions in the Understanding of Mucormycosis.Front Cell Infect Microbiol. 2022 Feb 24;12:850581. doi: 10.3389/fcimb.2022.850581. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35281441 Free PMC article. Review.
References
-
- Ben-Ami R. Angiogenesis at the mold-host interface: a potential key to understanding and treating invasive aspergillosis. Future Microbiol. 2013;8:1453–1462. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
