

Tumor treating fields perturb the localization of septins and cause aberrant mitotic exit
- PMID: 26010837
- PMCID: PMC4444126
- DOI: 10.1371/journal.pone.0125269
Tumor treating fields perturb the localization of septins and cause aberrant mitotic exit
Abstract
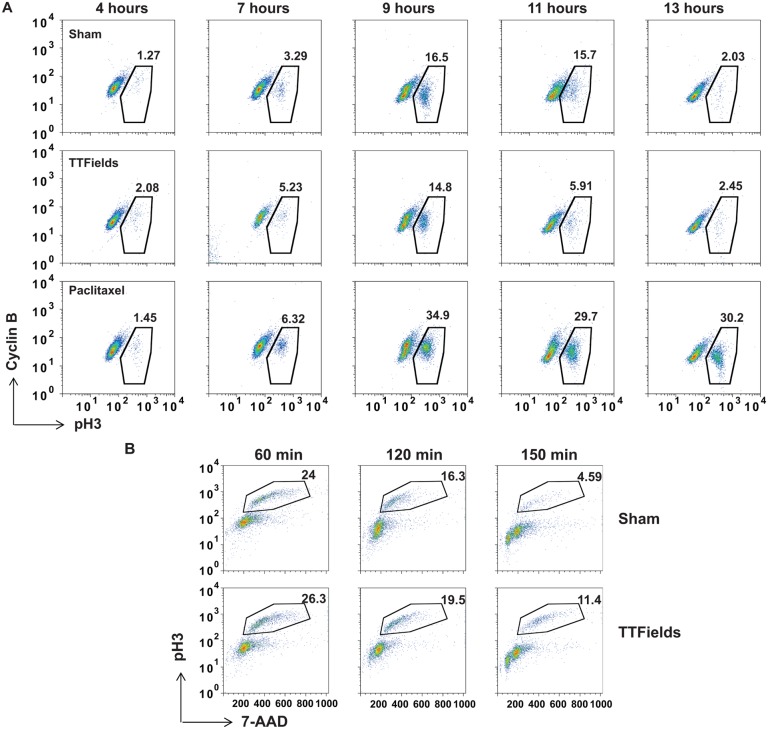
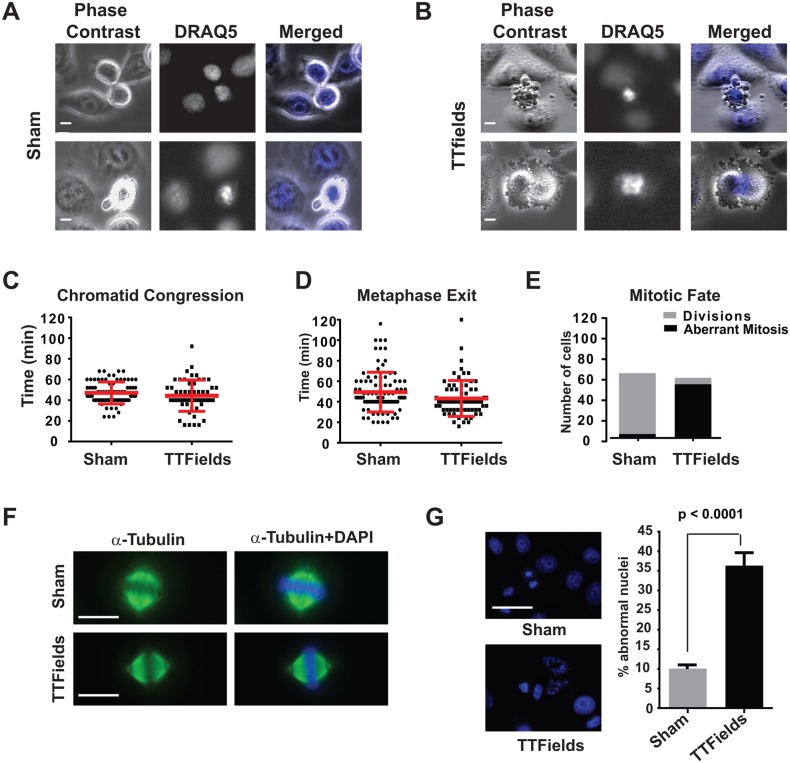
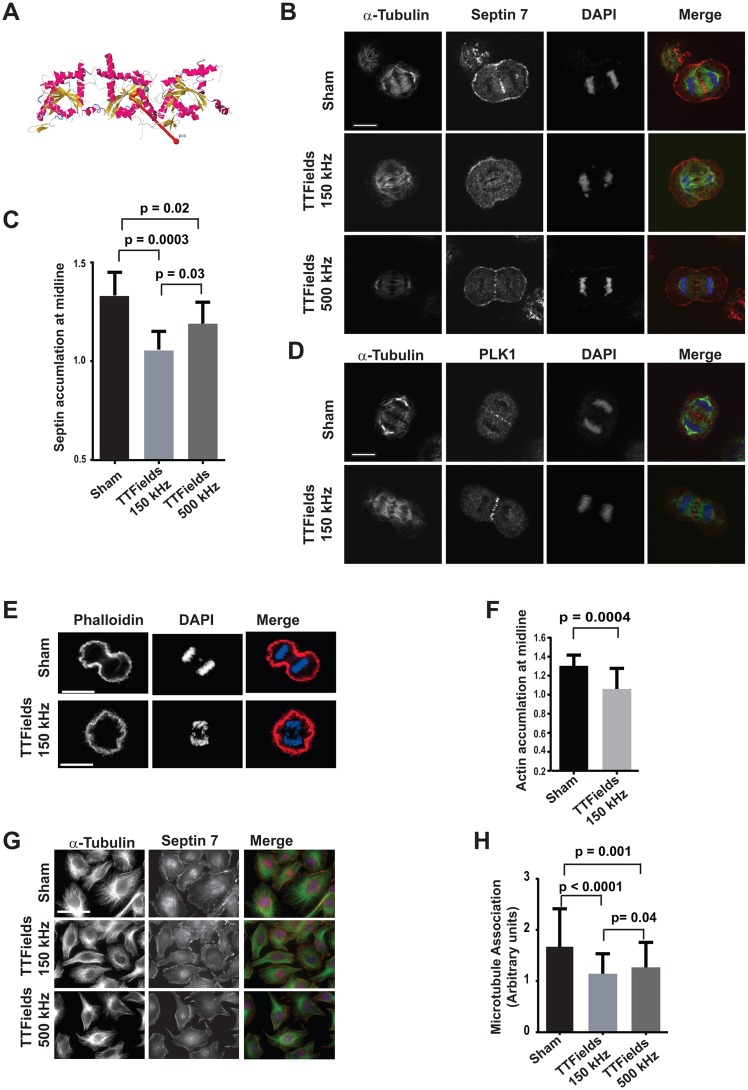
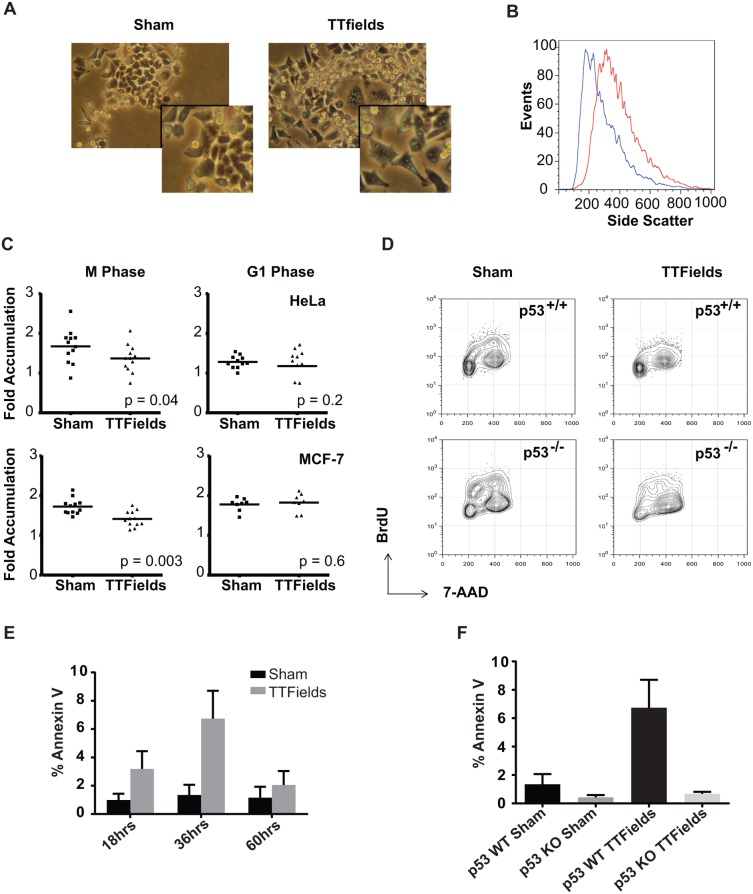
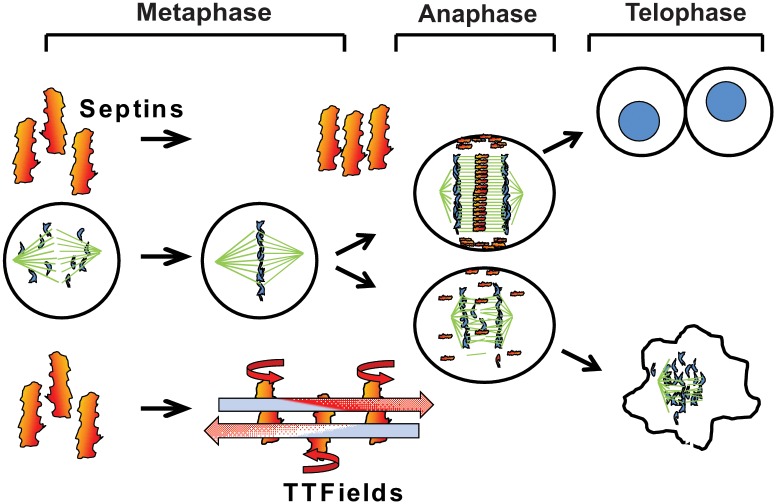
The anti-tumor effects of chemotherapy and radiation are thought to be mediated by triggering G1/S or G2/M cell cycle checkpoints, while spindle poisons, such as paclitaxel, block metaphase exit by initiating the spindle assembly checkpoint. In contrast, we have found that 150 kilohertz (kHz) alternating electric fields, also known as Tumor Treating Fields (TTFields), perturbed cells at the transition from metaphase to anaphase. Cells exposed to the TTFields during mitosis showed normal progression to this point, but exhibited uncontrolled membrane blebbing that coincided with metaphase exit. The ability of such alternating electric fields to affect cellular physiology is likely to be dependent on their interactions with proteins possessing high dipole moments. The mitotic Septin complex consisting of Septin 2, 6 and 7, possesses a high calculated dipole moment of 2711 Debyes (D) and plays a central role in positioning the cytokinetic cleavage furrow, and governing its contraction during ingression. We showed that during anaphase, TTFields inhibited Septin localization to the anaphase spindle midline and cytokinetic furrow, as well as its association with microtubules during cell attachment and spreading on fibronectin. After aberrant metaphase exit as a consequence of TTFields exposure, cells exhibited aberrant nuclear architecture and signs of cellular stress including an overall decrease in cellular proliferation, followed by apoptosis that was strongly influenced by the p53 mutational status. Thus, TTFields are able to diminish cell proliferation by specifically perturbing key proteins involved in cell division, leading to mitotic catastrophe and subsequent cell death.
Conflict of interest statement
Figures
References
-
- Gruber S, Haering CH, Nasmyth K. Chromosomal cohesin forms a ring. Cell. 2003;112(6):765–77. Epub 2003/03/26. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
