

Cofactor Specificity of the Bifunctional Alcohol and Aldehyde Dehydrogenase (AdhE) in Wild-Type and Mutant Clostridium thermocellum and Thermoanaerobacterium saccharolyticum
- PMID: 26013492
- PMCID: PMC4518838
- DOI: 10.1128/JB.00232-15
Cofactor Specificity of the Bifunctional Alcohol and Aldehyde Dehydrogenase (AdhE) in Wild-Type and Mutant Clostridium thermocellum and Thermoanaerobacterium saccharolyticum
Abstract
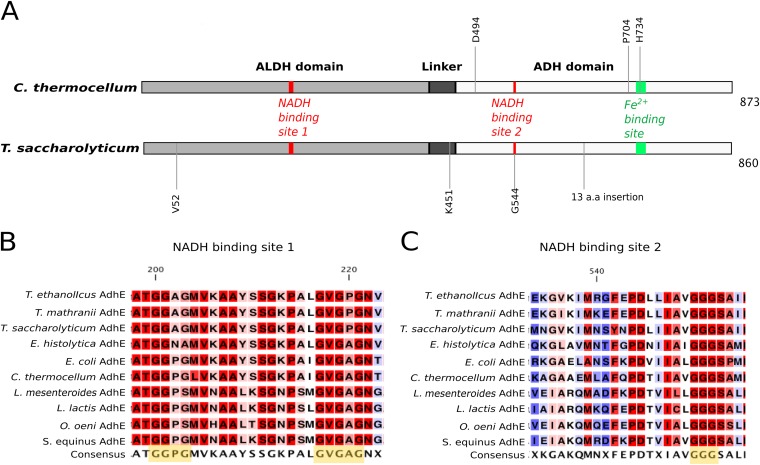
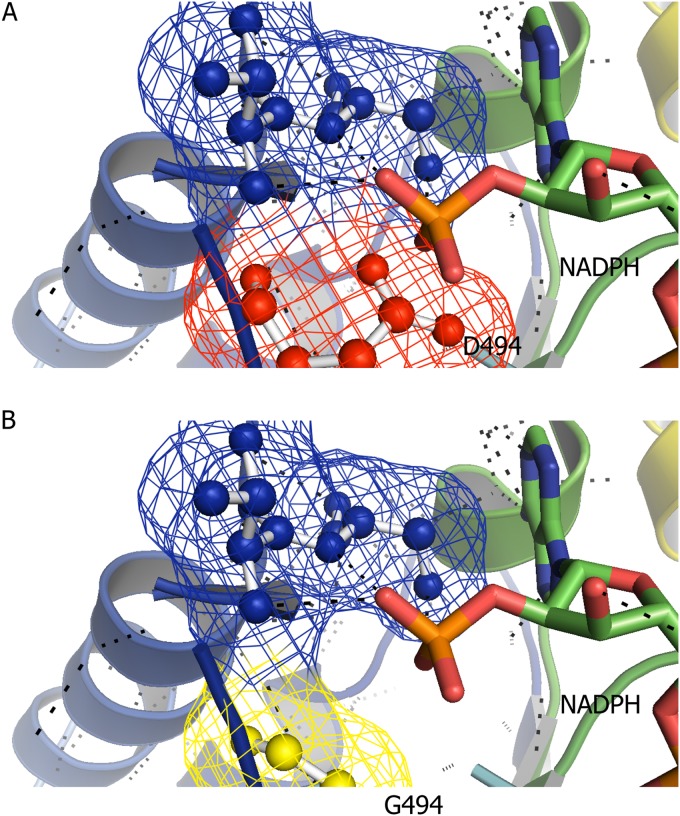
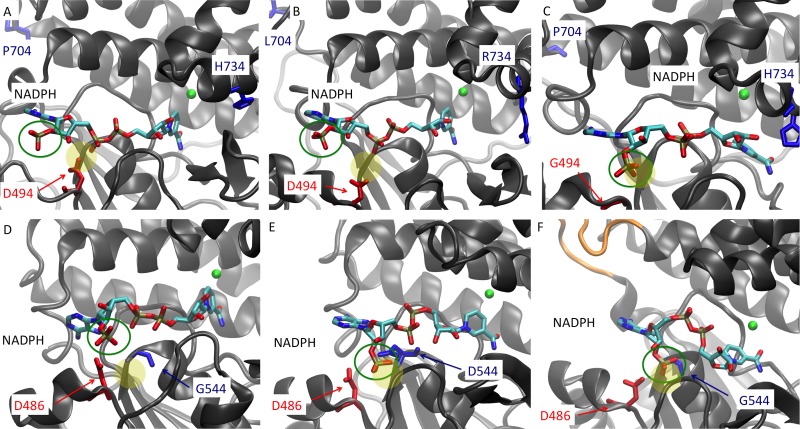
Clostridium thermocellum and Thermoanaerobacterium saccharolyticum are thermophilic bacteria that have been engineered to produce ethanol from the cellulose and hemicellulose fractions of biomass, respectively. Although engineered strains of T. saccharolyticum produce ethanol with a yield of 90% of the theoretical maximum, engineered strains of C. thermocellum produce ethanol at lower yields (∼50% of the theoretical maximum). In the course of engineering these strains, a number of mutations have been discovered in their adhE genes, which encode both alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) enzymes. To understand the effects of these mutations, the adhE genes from six strains of C. thermocellum and T. saccharolyticum were cloned and expressed in Escherichia coli, the enzymes produced were purified by affinity chromatography, and enzyme activity was measured. In wild-type strains of both organisms, NADH was the preferred cofactor for both ALDH and ADH activities. In high-ethanol-producing (ethanologen) strains of T. saccharolyticum, both ALDH and ADH activities showed increased NADPH-linked activity. Interestingly, the AdhE protein of the ethanologenic strain of C. thermocellum has acquired high NADPH-linked ADH activity while maintaining NADH-linked ALDH and ADH activities at wild-type levels. When single amino acid mutations in AdhE that caused increased NADPH-linked ADH activity were introduced into C. thermocellum and T. saccharolyticum, ethanol production increased in both organisms. Structural analysis of the wild-type and mutant AdhE proteins was performed to provide explanations for the cofactor specificity change on a molecular level.
Importance: This work describes the characterization of the AdhE enzyme from different strains of C. thermocellum and T. saccharolyticum. C. thermocellum and T. saccharolyticum are thermophilic anaerobes that have been engineered to make high yields of ethanol and can solubilize components of plant biomass and ferment the sugars to ethanol. In the course of engineering these strains, several mutations arose in the bifunctional ADH/ALDH protein AdhE, changing both enzyme activity and cofactor specificity. We show that changing AdhE cofactor specificity from mostly NADH linked to mostly NADPH linked resulted in higher ethanol production by C. thermocellum and T. saccharolyticum.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
The bifunctional alcohol and aldehyde dehydrogenase gene, adhE, is necessary for ethanol production in Clostridium thermocellum and Thermoanaerobacterium saccharolyticum.J Bacteriol. 2015 Apr;197(8):1386-93. doi: 10.1128/JB.02450-14. Epub 2015 Feb 9. J Bacteriol. 2015. PMID: 25666131 Free PMC article.
-
Characterization of the Clostridium thermocellum AdhE, NfnAB, ferredoxin and Pfor proteins for their ability to support high titer ethanol production in Thermoanaerobacterium saccharolyticum.Metab Eng. 2019 Jan;51:32-42. doi: 10.1016/j.ymben.2018.09.006. Epub 2018 Sep 12. Metab Eng. 2019. PMID: 30218716
-
The role of AdhE mutations in Thermoanaerobacterium saccharolyticum.J Bacteriol. 2025 May 22;207(5):e0001525. doi: 10.1128/jb.00015-25. Epub 2025 Apr 30. J Bacteriol. 2025. PMID: 40304514 Free PMC article.
-
Alcohol dehydrogenase: multiplicity and relatedness in the solvent-producing clostridia.FEMS Microbiol Rev. 1995 Oct;17(3):263-73. doi: 10.1111/j.1574-6976.1995.tb00210.x. FEMS Microbiol Rev. 1995. PMID: 7576768 Review.
-
Alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) in the cancer diseases.Clin Chim Acta. 2008 Sep;395(1-2):1-5. doi: 10.1016/j.cca.2008.05.001. Epub 2008 May 8. Clin Chim Acta. 2008. PMID: 18505683 Review.
Cited by
-
Determining the roles of the three alcohol dehydrogenases (AdhA, AdhB and AdhE) in Thermoanaerobacter ethanolicus during ethanol formation.J Ind Microbiol Biotechnol. 2017 May;44(4-5):745-757. doi: 10.1007/s10295-016-1896-6. Epub 2017 Jan 11. J Ind Microbiol Biotechnol. 2017. PMID: 28078513
-
Deletion of the hfsB gene increases ethanol production in Thermoanaerobacterium saccharolyticum and several other thermophilic anaerobic bacteria.Biotechnol Biofuels. 2017 Nov 30;10:282. doi: 10.1186/s13068-017-0968-9. eCollection 2017. Biotechnol Biofuels. 2017. PMID: 29213322 Free PMC article.
-
A mutation in the AdhE alcohol dehydrogenase of Clostridium thermocellum increases tolerance to several primary alcohols, including isobutanol, n-butanol and ethanol.Sci Rep. 2019 Feb 11;9(1):1736. doi: 10.1038/s41598-018-37979-5. Sci Rep. 2019. PMID: 30741948 Free PMC article.
-
Targeted redox and energy cofactor metabolomics in Clostridium thermocellum and Thermoanaerobacterium saccharolyticum.Biotechnol Biofuels. 2017 Nov 30;10:270. doi: 10.1186/s13068-017-0960-4. eCollection 2017. Biotechnol Biofuels. 2017. PMID: 29213318 Free PMC article.
-
Deletion of nfnAB in Thermoanaerobacterium saccharolyticum and Its Effect on Metabolism.J Bacteriol. 2015 Sep;197(18):2920-9. doi: 10.1128/JB.00347-15. Epub 2015 Jun 29. J Bacteriol. 2015. PMID: 26124241 Free PMC article.
References
-
- Mai V, Lorenz WW, Wiegel J. 1997. Transformation of Thermoanaerobacterium sp. strain JW/SL-YS485 with plasmid pIKM1 conferring kanamycin resistance. FEMS Microbiol Lett 148:163–167. doi:10.1111/j.1574-6968.1997.tb10283.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
