

Niemann-pick C1 is essential for ebolavirus replication and pathogenesis in vivo
- PMID: 26015498
- PMCID: PMC4447246
- DOI: 10.1128/mBio.00565-15
Niemann-pick C1 is essential for ebolavirus replication and pathogenesis in vivo
Abstract
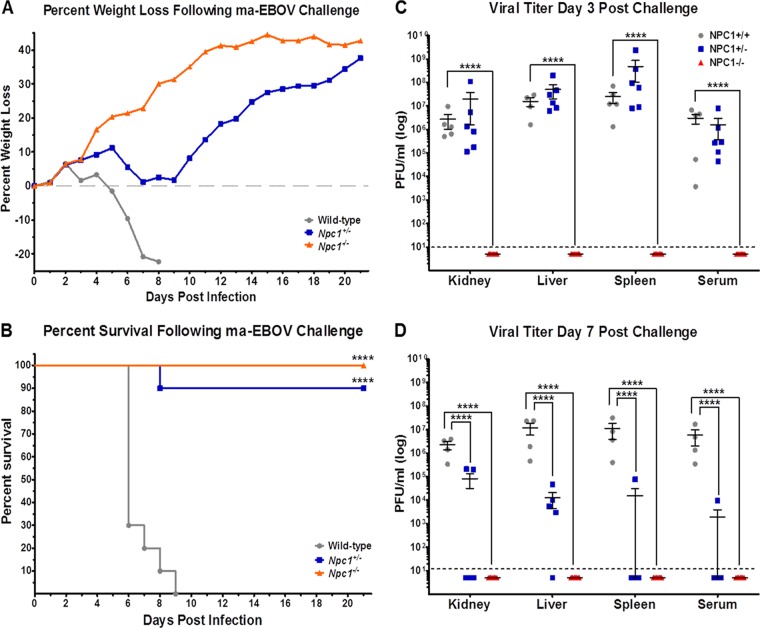
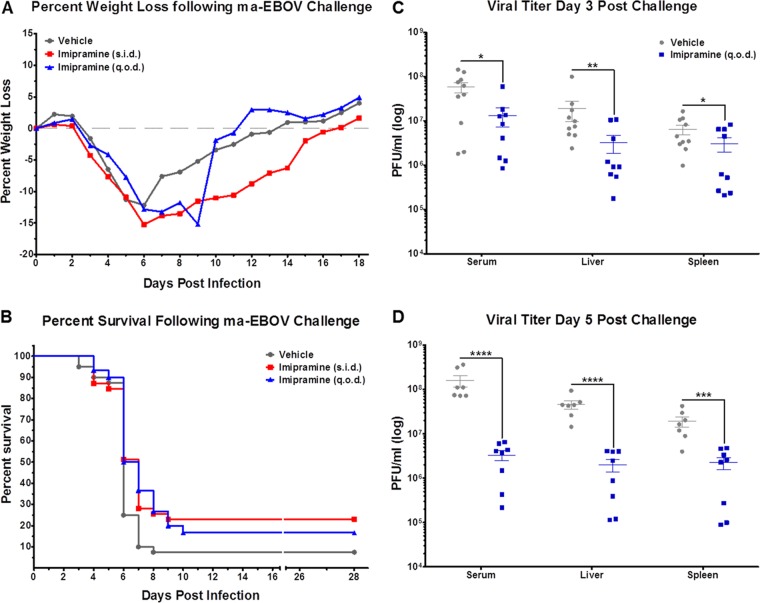
Recent work demonstrated that the Niemann-Pick C1 (NPC1) protein is an essential entry receptor for filoviruses. While previous studies focused on filovirus entry requirements of NPC1 in vitro, its roles in filovirus replication and pathogenesis in vivo remain unclear. Here, we evaluated the importance of NPC1, and its partner in cholesterol transport, NPC2, by using a mouse model of Ebolavirus (EBOV) disease. We found that, whereas wild-type mice had high viral loads and succumbed to EBOV infection, Npc1(-/-) mice were entirely free of viral replication and completely protected from EBOV disease. Interestingly, Npc1(+/-) mice transiently developed high levels of viremia, but were nevertheless substantially protected from EBOV challenge. We also found Npc2(-/-) mice to be fully susceptible to EBOV infection, while Npc1(-/-) mice treated to deplete stored lysosomal cholesterol remained completely resistant to EBOV infection. These results provide mechanistic evidence that NPC1 is directly required for EBOV infection in vivo, with little or no role for NPC1/NPC2-dependent cholesterol transport. Finally, we assessed the in vivo antiviral efficacies of three compounds known to inhibit NPC1 function or NPC1-glycoprotein binding in vitro. Two compounds reduced viral titers in vivo and provided a modest, albeit not statistically significant, degree of protection. Taken together, our results show that NPC1 is critical for replication and pathogenesis in animals and is a bona fide target for development of antifilovirus therapeutics. Additionally, our findings with Npc1(+/-) mice raise the possibility that individuals heterozygous for NPC1 may have a survival advantage in the face of EBOV infection.
Importance: Researchers have been searching for an essential filovirus receptor for decades, and numerous candidate receptors have been proposed. However, none of the proposed candidate receptors has proven essential in all in vitro scenarios, nor have they proven essential when evaluated using animal models. In this report, we provide the first example of a knockout mouse that is completely refractory to EBOV infection, replication, and disease. The findings detailed here provide the first critical in vivo data illustrating the absolute requirement of NPC1 for filovirus infection in mice. Our work establishes NPC1 as a legitimate target for the development of anti-EBOV therapeutics. However, the limited success of available NPC1 inhibitors to protect mice from EBOV challenge highlights the need for new molecules or approaches to target NPC1 in vivo.
Copyright © 2015 Herbert et al.
Figures
References
-
- Kuhn JH, Becker S, Ebihara H, Geisbert TW, Johnson KM, Kawaoka Y, Lipkin WI, Negredo AI, Netesov SV, Nichol ST, Palacios G, Peters CJ, Tenorio A, Volchkov VE, Jahrling PB. 2010. Proposal for a revised taxonomy of the family Filoviridae: classification, names of taxa and viruses, and virus abbreviations. Arch Virol 155:2083–2103. doi:10.1007/s00705-010-0814-x. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
