

The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics
- PMID: 26015579
- PMCID: PMC4466716
- DOI: 10.1073/pnas.1504822112
The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics
Abstract
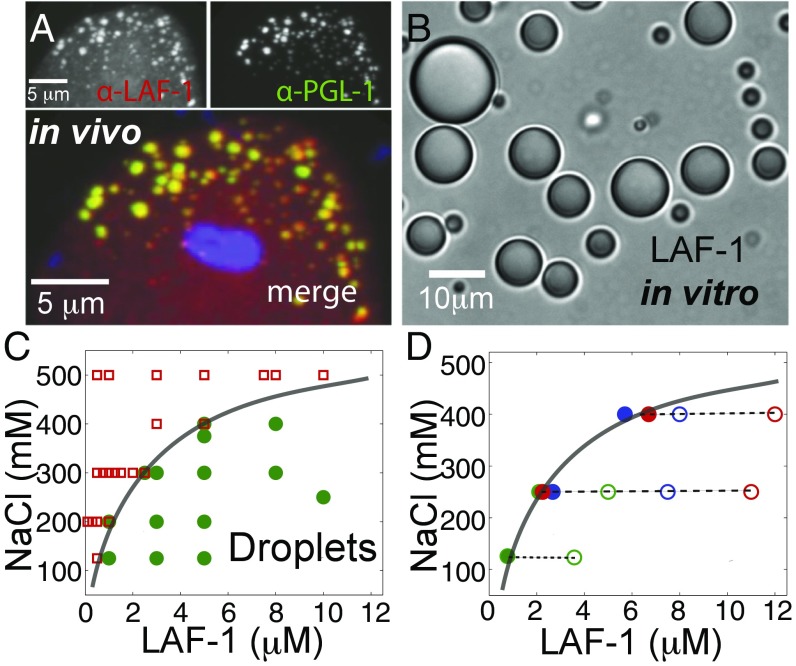
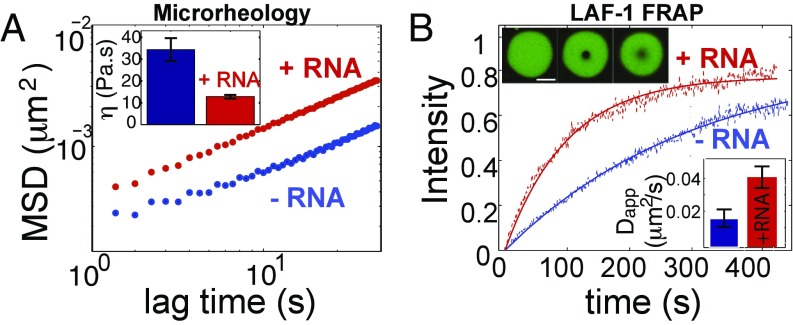
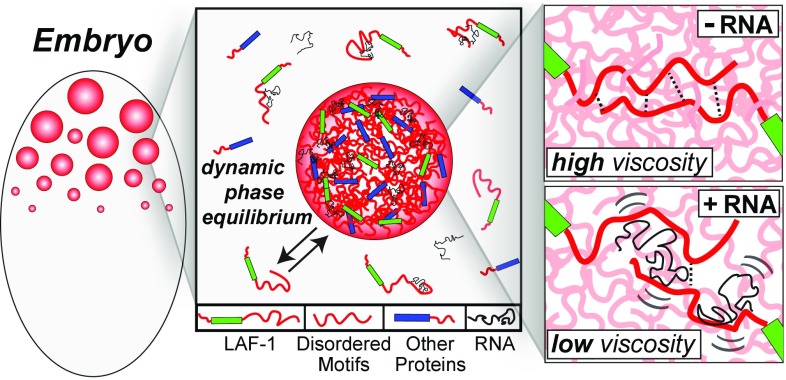
P granules and other RNA/protein bodies are membrane-less organelles that may assemble by intracellular phase separation, similar to the condensation of water vapor into droplets. However, the molecular driving forces and the nature of the condensed phases remain poorly understood. Here, we show that the Caenorhabditis elegans protein LAF-1, a DDX3 RNA helicase found in P granules, phase separates into P granule-like droplets in vitro. We adapt a microrheology technique to precisely measure the viscoelasticity of micrometer-sized LAF-1 droplets, revealing purely viscous properties highly tunable by salt and RNA concentration. RNA decreases viscosity and increases molecular dynamics within the droplet. Single molecule FRET assays suggest that this RNA fluidization results from highly dynamic RNA-protein interactions that emerge close to the droplet phase boundary. We demonstrate than an N-terminal, arginine/glycine rich, intrinsically disordered protein (IDP) domain of LAF-1 is necessary and sufficient for both phase separation and RNA-protein interactions. In vivo, RNAi knockdown of LAF-1 results in the dissolution of P granules in the early embryo, with an apparent submicromolar phase boundary comparable to that measured in vitro. Together, these findings demonstrate that LAF-1 is important for promoting P granule assembly and provide insight into the mechanism by which IDP-driven molecular interactions give rise to liquid phase organelles with tunable properties.
Keywords: RNA granules; intracellular phase transition; intrinsically disordered proteins; liquid droplets.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Brangwynne CP, et al. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science. 2009;324(5935):1729–1732. - PubMed
-
- Wippich F, et al. Dual specificity kinase DYRK3 couples stress granule condensation/dissolution to mTORC1 signaling. Cell. 2013;152(4):791–805. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
