

Complementary expression and phosphorylation of Cx46 and Cx50 during development and following gene deletion in mouse and in normal and orchitic mink testes
- PMID: 26017495
- PMCID: PMC4525330
- DOI: 10.1152/ajpregu.00152.2015
Complementary expression and phosphorylation of Cx46 and Cx50 during development and following gene deletion in mouse and in normal and orchitic mink testes
Erratum in
-
Corrigendum.Am J Physiol Regul Integr Comp Physiol. 2015 Dec 15;309(12):R1578. doi: 10.1152/ajpregu.zh6-8905-corr.2015. Am J Physiol Regul Integr Comp Physiol. 2015. PMID: 26671929 Free PMC article. No abstract available.
Abstract
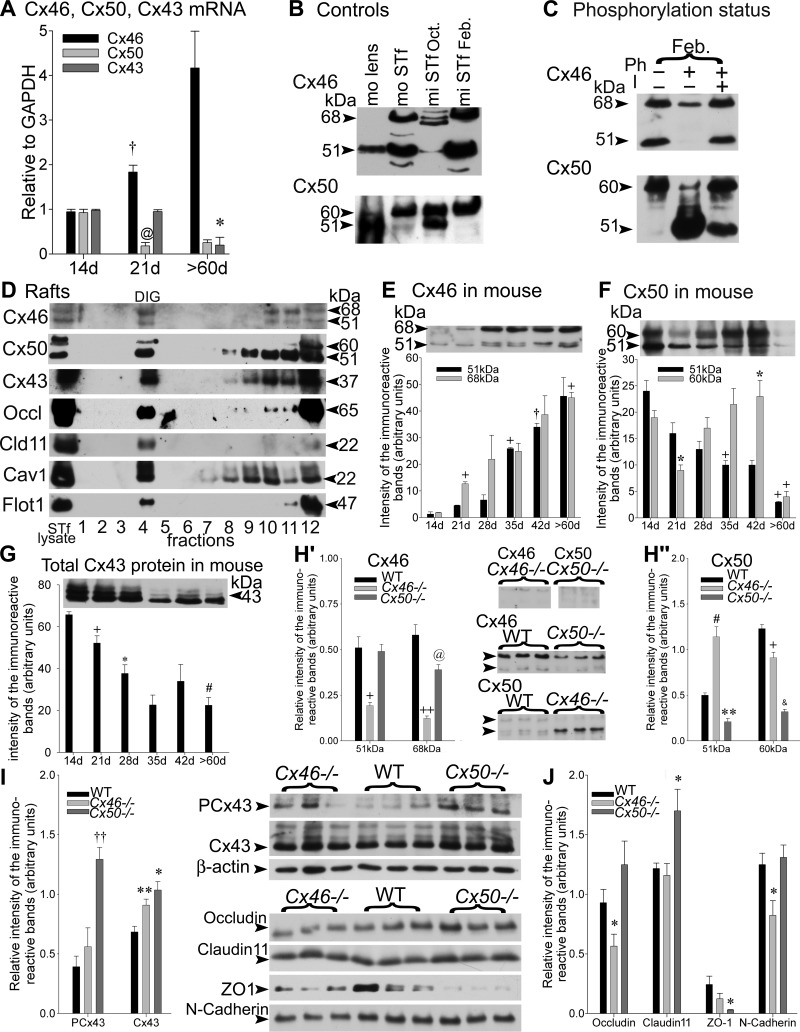
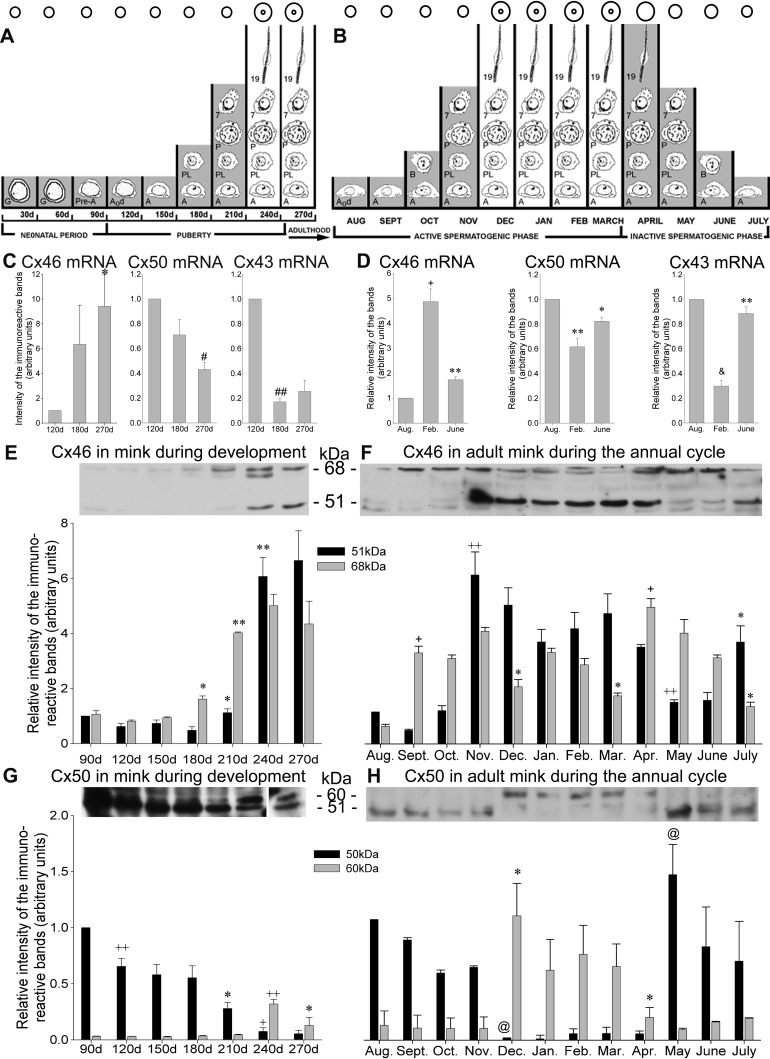
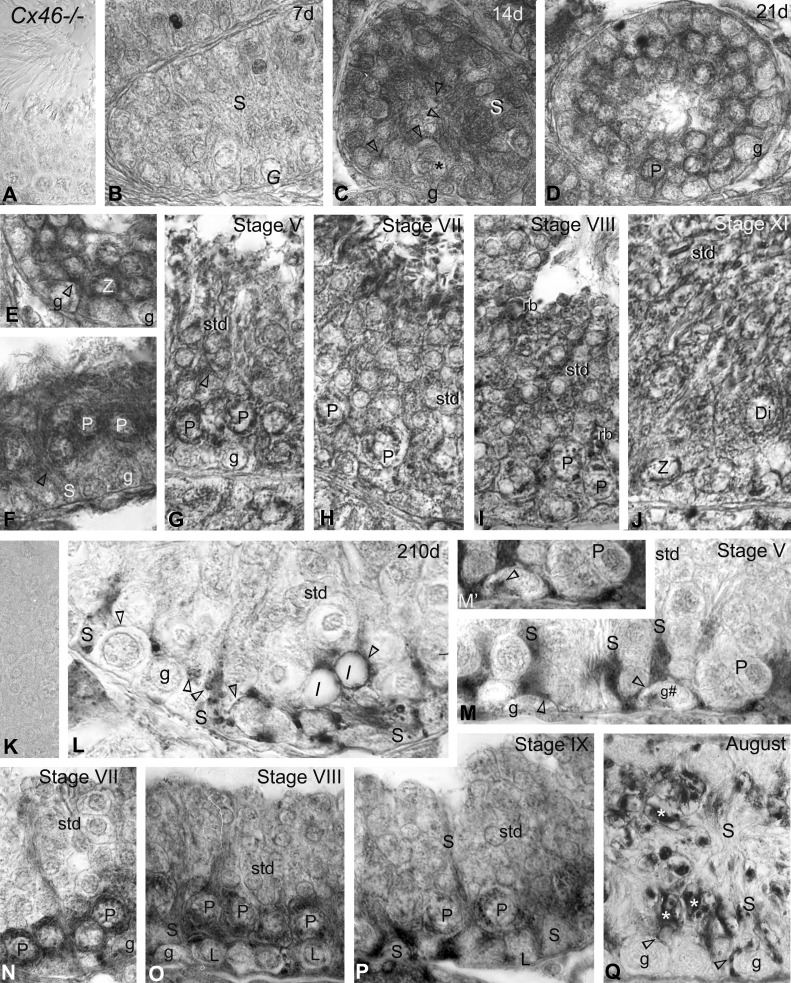
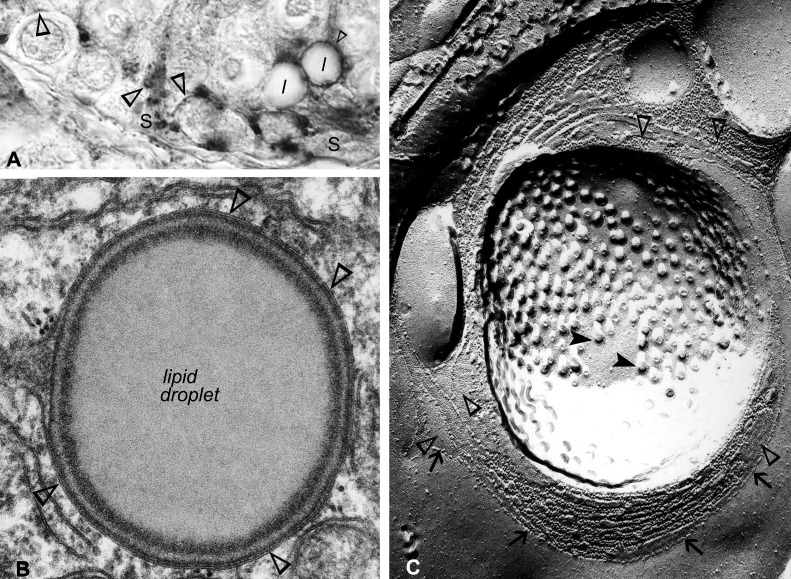
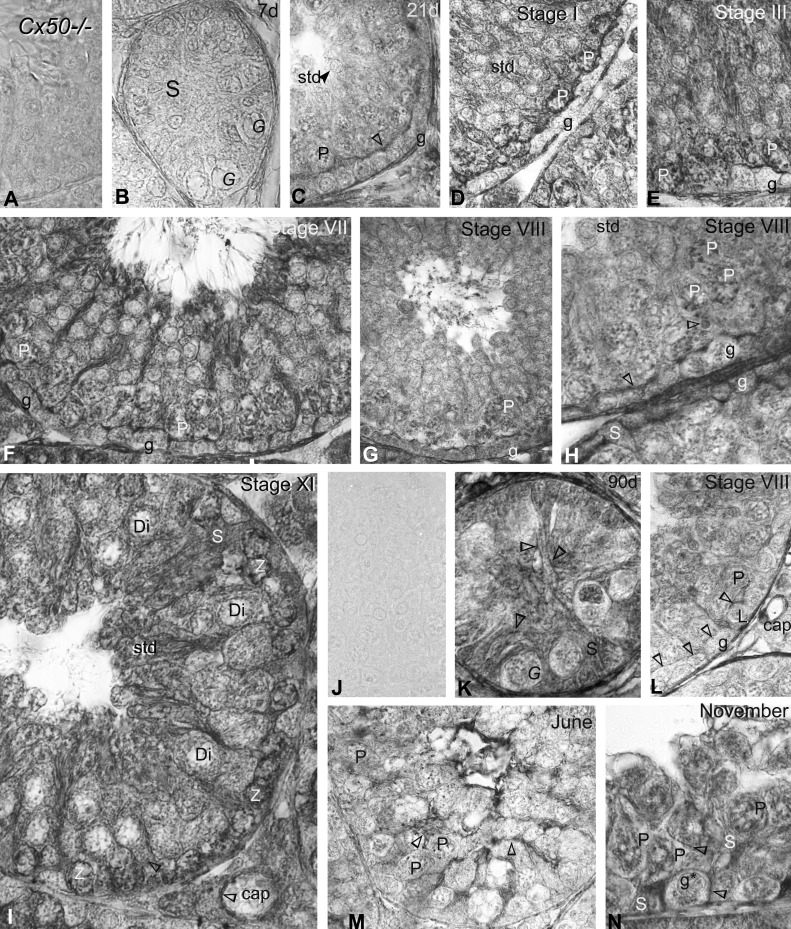
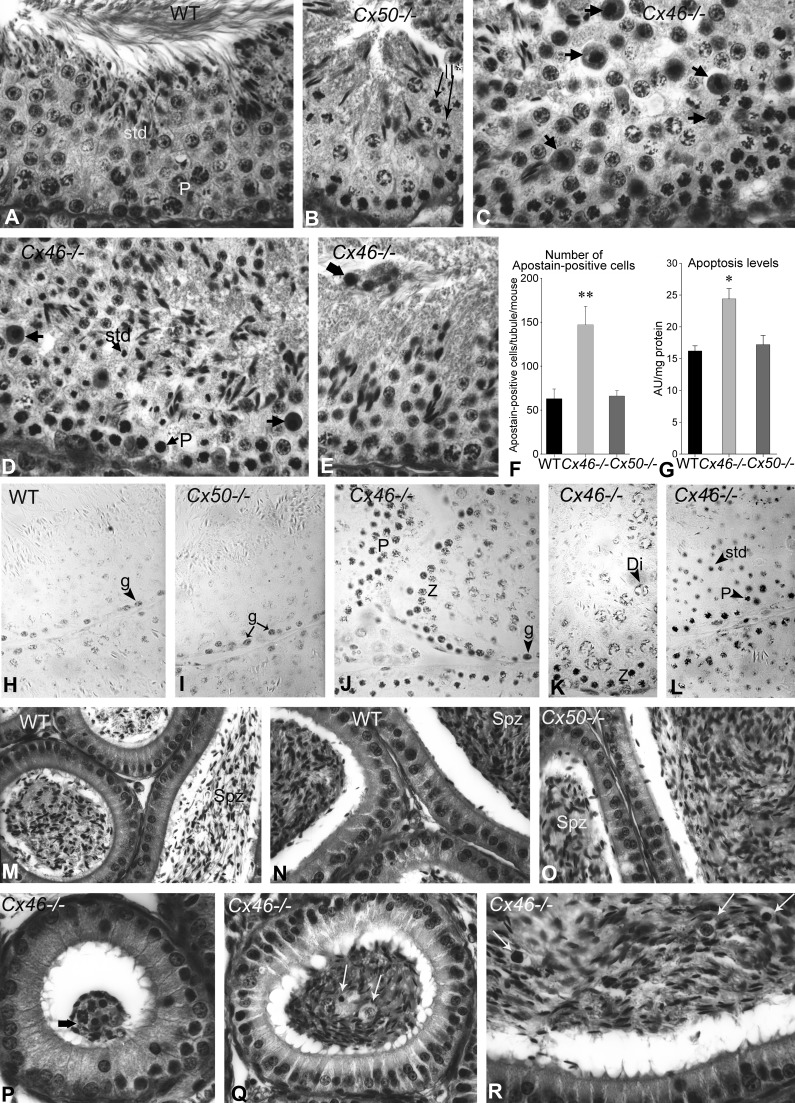
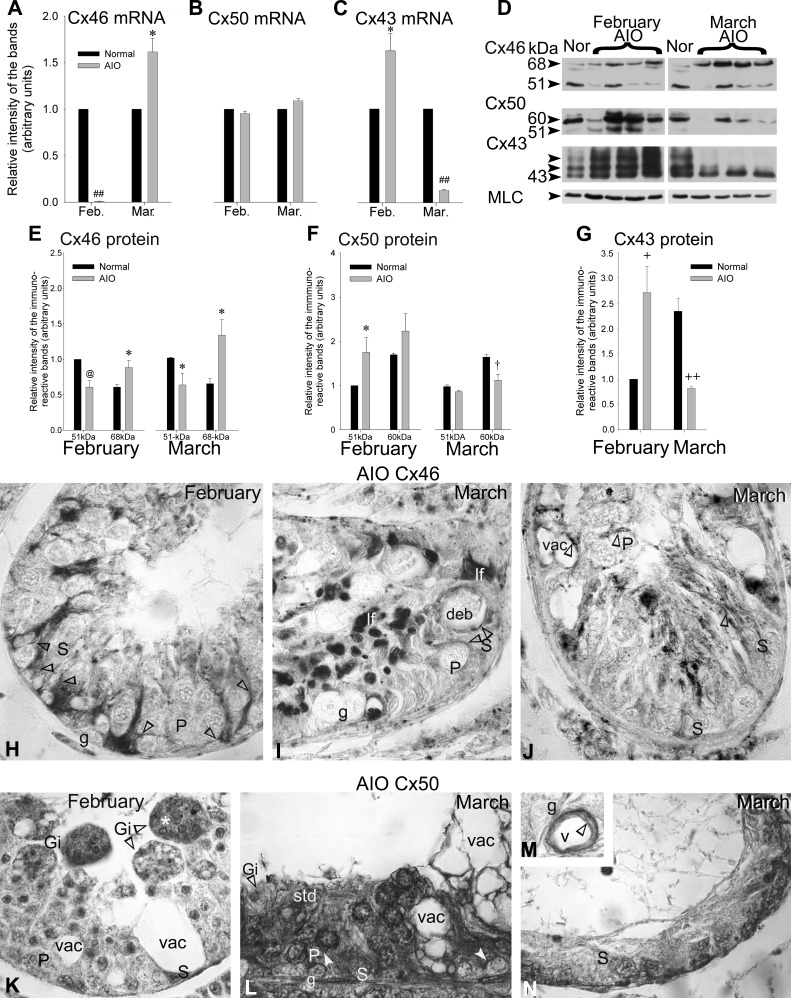
Gap junction-mediated communication helps synchronize interconnected Sertoli cell activities. Besides, coordination of germ cell and Sertoli cell activities depends on gap junction-mediated Sertoli cell-germ cell communication. This report assesses mechanisms underlying the regulation of connexin 46 (Cx46) and Cx50 in mouse testis and those accompanying a "natural" seasonal and a pathological arrest of spermatogenesis, resulting from autoimmune orchitis (AIO) in mink. Furthermore, the impact of deleting Cx46 or Cx50 on the expression, phosphorylation of junction proteins, and spermatogenesis is evaluated. Cx46 mRNA and protein expression increased, whereas Cx50 decreased with adulthood in normal mice and mink. Cx46 mRNA and protein expression increased, whereas Cx50 decreased with adulthood in normal mice and mink. During the mink active spermatogenic phase, Cx50 became phosphorylated and localized to the site of the blood-testis barrier. By contrast, Cx46 was dephosphorylated and associated with annular junctions, suggesting phosphorylation/dephosphorylation of Cx46 and Cx50 involvement in the barrier dynamics. Cx46-positive annular junctions in contact with lipid droplets were found. Cx46 and Cx50 expression and localization were altered in mink with AIO. The deletion of Cx46 or Cx50 impacted on other connexin expression and phosphorylation and differently affected tight and adhering junction protein expression. The level of apoptosis, determined by ELISA, and a number of Apostain-labeled spermatocytes and spermatids/tubules were higher in mice lacking Cx46 (Cx46-/-) than wild-type and Cx50-/- mice, arguing for life-sustaining Cx46 gap junction-mediated exchanges in late-stage germ cells secluded from the blood by the barrier. The data show that expression and phosphorylation of Cx46 and Cx50 are complementary in seminiferous tubules.
Keywords: Sertoli cell; connexin 43; lipid droplet; meiosis; seasonal breeder.
Copyright © 2015 the American Physiological Society.
Figures
Similar articles
-
CX43 expression, phosphorylation, and distribution in the normal and autoimmune orchitic testis with a look at gap junctions joining germ cell to germ cell.Am J Physiol Regul Integr Comp Physiol. 2011 Jan;300(1):R121-39. doi: 10.1152/ajpregu.00500.2010. Epub 2010 Oct 20. Am J Physiol Regul Integr Comp Physiol. 2011. PMID: 20962206
-
Distinctive actions of connexin 46 and connexin 50 in anterior pituitary folliculostellate cells.PLoS One. 2017 Jul 31;12(7):e0182495. doi: 10.1371/journal.pone.0182495. eCollection 2017. PLoS One. 2017. PMID: 28759642 Free PMC article.
-
Cx30.2 deletion causes imbalances in testicular Cx43, Cx46, and Cx50 and insulin receptors. Reciprocally, diabetes/obesity alters Cx30.2 in mouse testis.Am J Physiol Regul Integr Comp Physiol. 2020 Jun 1;318(6):R1078-R1090. doi: 10.1152/ajpregu.00044.2020. Epub 2020 Apr 29. Am J Physiol Regul Integr Comp Physiol. 2020. PMID: 32348681 Free PMC article.
-
Gap junctions or hemichannel-dependent and independent roles of connexins in cataractogenesis and lens development.Curr Mol Med. 2010 Dec;10(9):851-63. doi: 10.2174/156652410793937750. Curr Mol Med. 2010. PMID: 21091421 Free PMC article. Review.
-
Mutations of CX46/CX50 and Cataract Development.Front Mol Biosci. 2022 Feb 11;9:842399. doi: 10.3389/fmolb.2022.842399. eCollection 2022. Front Mol Biosci. 2022. PMID: 35223995 Free PMC article. Review.
Cited by
-
Exploring the role of endogenous retroviruses in seasonal reproductive cycles: a case study of the ERV-V envelope gene in mink.Front Cell Infect Microbiol. 2024 Jun 28;14:1404431. doi: 10.3389/fcimb.2024.1404431. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 39081866 Free PMC article.
-
Glucose, insulin, insulin receptor subunits α and β in normal and spontaneously diabetic and obese ob/ob and db/db infertile mouse testis and hypophysis.Reprod Biol Endocrinol. 2020 Mar 17;18(1):25. doi: 10.1186/s12958-020-00583-2. Reprod Biol Endocrinol. 2020. PMID: 32183843 Free PMC article.
-
Action and Interaction between Retinoic Acid Signaling and Blood-Testis Barrier Function in the Spermatogenesis Cycle.Cells. 2022 Jan 21;11(3):352. doi: 10.3390/cells11030352. Cells. 2022. PMID: 35159162 Free PMC article. Review.
-
What do we know about blood-testis barrier? current understanding of its structure and physiology.Front Cell Dev Biol. 2023 Jun 15;11:1114769. doi: 10.3389/fcell.2023.1114769. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37397257 Free PMC article. Review.
-
PCSK9 Contributes to the Cholesterol, Glucose, and Insulin2 Homeostasis in Seminiferous Tubules and Maintenance of Immunotolerance in Testis.Front Cell Dev Biol. 2022 May 2;10:889972. doi: 10.3389/fcell.2022.889972. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35586340 Free PMC article.
References
-
- Abraham V, Chou ML, George P, Pooler P, Zaman A, Savani R, Koval M. Heterocellular gap junctional communication between alveolar epithelial cells. Am J Physiol Lung Cell Mol Physiol 280: L1085–L1093, 2001. - PubMed
-
- Akpovi CD, Pelletier RM. A revised and improved method for the isolation of seminiferous tubule-enriched fractions that preserves the phosphorylated and glycosylated forms of proteins. In: Human Embryogenesis Methods and Protocols, edited by Lafond J, Vaillancourt C. Totowa, NJ: Humana, Springer Protocols, 2009, p. 159–168. - PubMed
-
- Akpovi CD, Yoon SR, Vitale ML, Pelletier RM. The predominance of one of the SR-BI isoforms is associated with increased esterified cholesterol levels not apoptosis in mink testis. J Lipid Res 47: 2233–2247, 2006. - PubMed
-
- Alcala J, Katar M, Maisel H. Lipid composition of chick lens fiber cell gap junctions. Curr Eye Res 2: 569–578, 1982. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous