

CXCL12 Signaling Is Essential for Maturation of the Ventricular Coronary Endothelial Plexus and Establishment of Functional Coronary Circulation
- PMID: 26017771
- PMCID: PMC4448078
- DOI: 10.1016/j.devcel.2015.03.018
CXCL12 Signaling Is Essential for Maturation of the Ventricular Coronary Endothelial Plexus and Establishment of Functional Coronary Circulation
Abstract
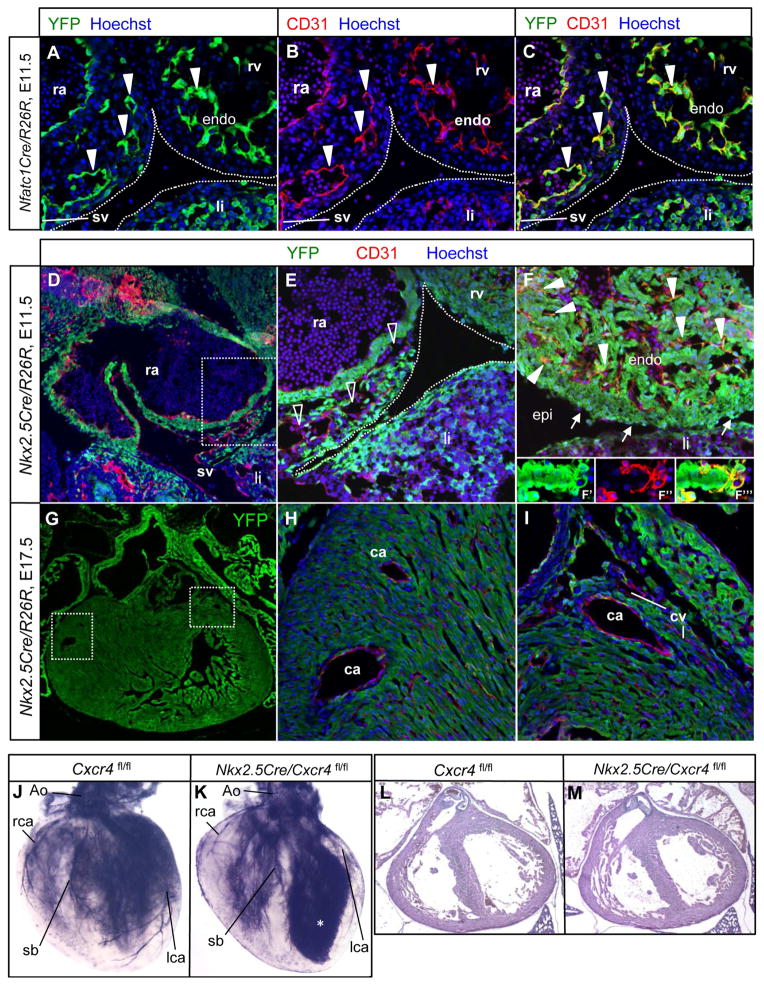
Maturation of a vascular plexus is a critical and yet incompletely understood process in organ development, and known maturation factors act universally in all vascular beds. In this study, we show that CXCL12 is an organ-specific maturation factor of particular relevance in coronary arterial vasculature. In vitro, CXCL12 does not influence nascent vessel formation, but promotes higher-order complexity of preinitiated vessels. In the heart, CXCL12 is expressed principally by the epicardium, and its receptor CXCR4 is expressed by coronary endothelial cells. CXCL12 is not a chemotactic signal for endothelial cell migration, but rather acts in a paracrine manner to influence the maturation of the coronary vascular plexus. Mutants in CXCL12 signaling show an excess of immature capillary chains and a selective failure in arterial maturation, and become leaky with the onset of coronary perfusion. Failed maturation of the coronary system explains the late-gestation lethality of these mutants.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures



References
-
- Ara T, Tokoyoda K, Okamoto R, Koni PA, Nagasawa T. The role of CXCL12 in the organ-specific process of artery formation. Blood. 2005;105:3155–3161. - PubMed
-
- Carmeliet P, Ferreira V, Breier G, Pollefeyt S, Kieckens L, Gertsenstein M, Fahrig M, Vandenhoeck A, Harpal K, Eberhardt C, et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature. 1996;380:435–439. - PubMed
-
- Choudhary B, Zhou J, Li P, Thomas S, Kaartinen V, Sucov HM. Absence of TGFbeta signaling in embryonic vascular smooth muscle leads to reduced lysyl oxidase expression, impaired elastogenesis, and aneurysm. Genesis. 2009;47:115–121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
