

The sensory ecology of adaptive landscapes
- PMID: 26018831
- PMCID: PMC4455732
- DOI: 10.1098/rsbl.2014.1054
The sensory ecology of adaptive landscapes
Abstract
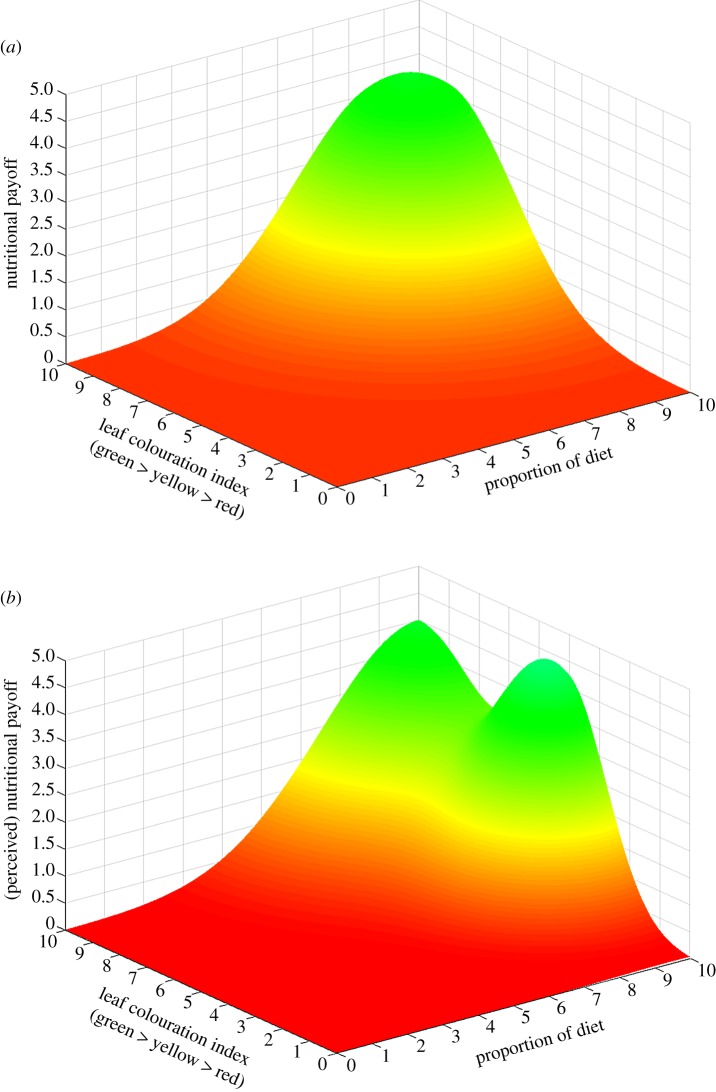
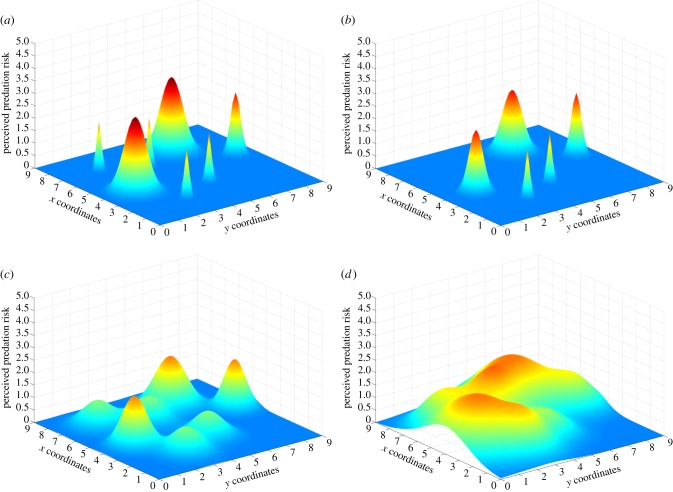
In complex environments, behavioural plasticity depends on the ability of an animal to integrate numerous sensory stimuli. The multidimensionality of factors interacting to shape plastic behaviour means it is difficult for both organisms and researchers to predict what constitutes an adaptive response to a given set of conditions. Although researchers may be able to map the fitness pay-offs of different behavioural strategies in changing environments, there is no guarantee that the study species will be able to perceive these pay-offs. We thus risk a disconnect between our own predictions about adaptive behaviour and what is behaviourally achievable given the umwelt of the animal being studied. This may lead to erroneous conclusions about maladaptive behaviour in circumstances when the behaviour exhibited is the most adaptive possible given sensory limitations. With advances in the computational resources available to behavioural ecologists, we can now measure vast numbers of interactions among behaviours and environments to create adaptive behavioural surfaces. These surfaces have massive heuristic, predictive and analytical potential in understanding adaptive animal behaviour, but researchers using them are destined to fail if they ignore the sensory ecology of the species they study. Here, we advocate the continued use of these approaches while directly linking them to perceptual space to ensure that the topology of the generated adaptive landscape matches the perceptual reality of the animal it intends to study. Doing so will allow predictive models of animal behaviour to reflect the reality faced by the agents on adaptive surfaces, vastly improving our ability to determine what constitutes an adaptive response for the animal in question.
Keywords: adaptive landscape; behavioural plasticity; fitness surface; perception; sensory ecology.
© 2015 The Author(s) Published by the Royal Society. All rights reserved.
Figures
References
-
- DeWitt TJ, Scheiner SM. 2004. Phenotypic plasticity: functional and conceptual approaches. Oxford, UK: Oxford University Press.
-
- West-Eberhard MJ. 2003. Developmental plasticity and evolution. Oxford, UK: Oxford University Press.
-
- Ghalambor CK, Angeloni LM, Carroll SP. 2010. Behavior as phenotypic plasticity. In Evolutionary behavioral ecology (eds Westneat DF, Fox CW.), pp. 90–107. Oxford, UK: Oxford University Press.
-
- Dusenbery DB. 1992. Sensory ecology: how organisms acquire and respond to information. New York, NY: WH Freeman.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources