

Multiscale mechanobiology: computational models for integrating molecules to multicellular systems
- PMID: 26019013
- PMCID: PMC4677065
- DOI: 10.1039/c5ib00043b
Multiscale mechanobiology: computational models for integrating molecules to multicellular systems
Abstract
Mechanical signals exist throughout the biological landscape. Across all scales, these signals, in the form of force, stiffness, and deformations, are generated and processed, resulting in an active mechanobiological circuit that controls many fundamental aspects of life, from protein unfolding and cytoskeletal remodeling to collective cell motions. The multiple scales and complex feedback involved present a challenge for fully understanding the nature of this circuit, particularly in development and disease in which it has been implicated. Computational models that accurately predict and are based on experimental data enable a means to integrate basic principles and explore fine details of mechanosensing and mechanotransduction in and across all levels of biological systems. Here we review recent advances in these models along with supporting and emerging experimental findings.
Figures
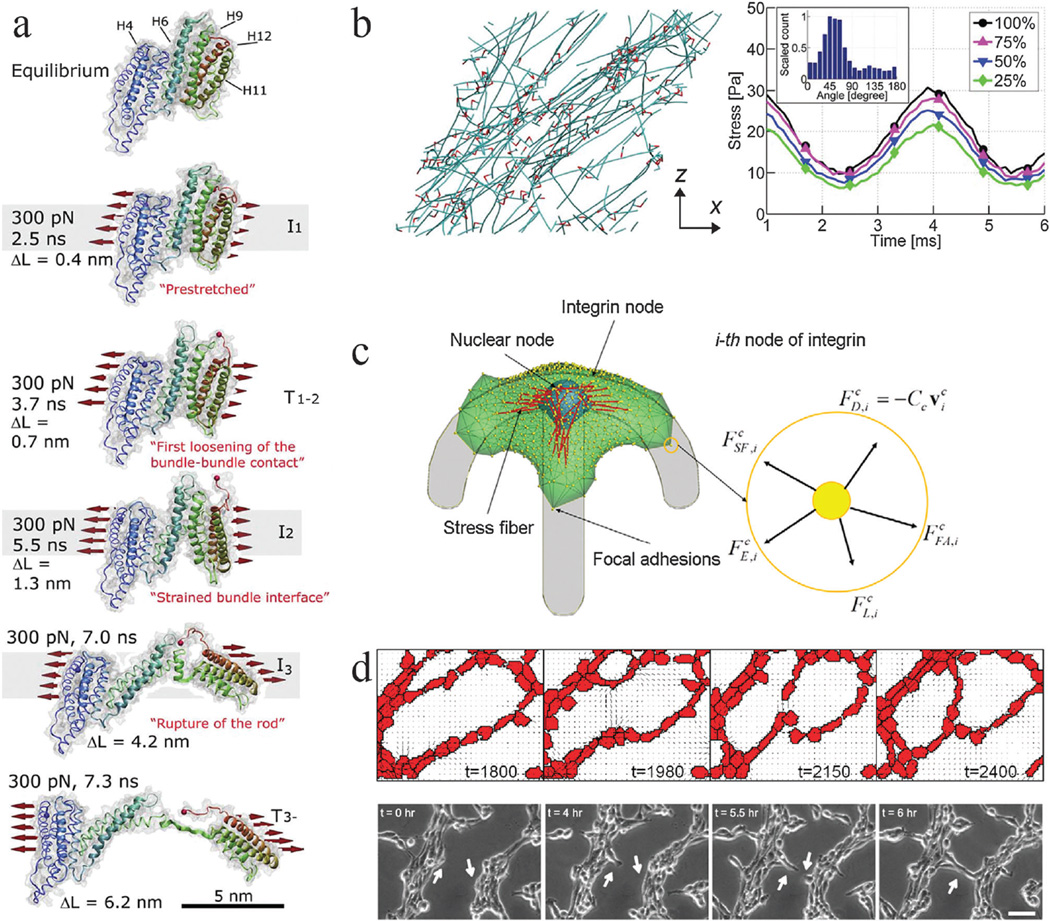
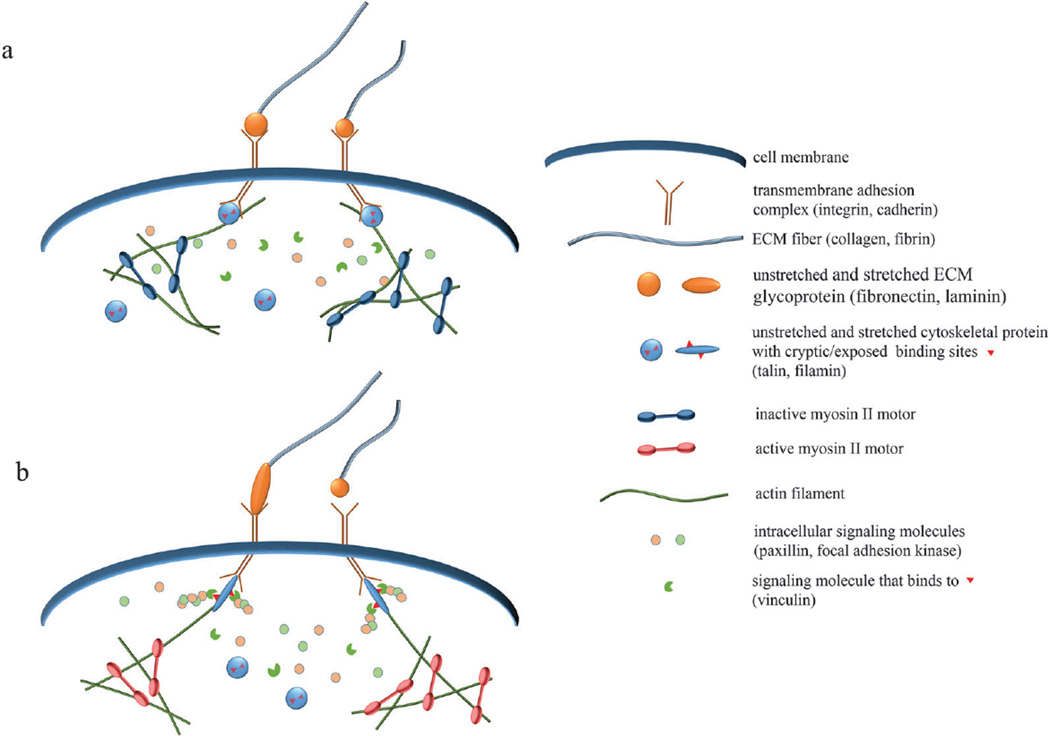
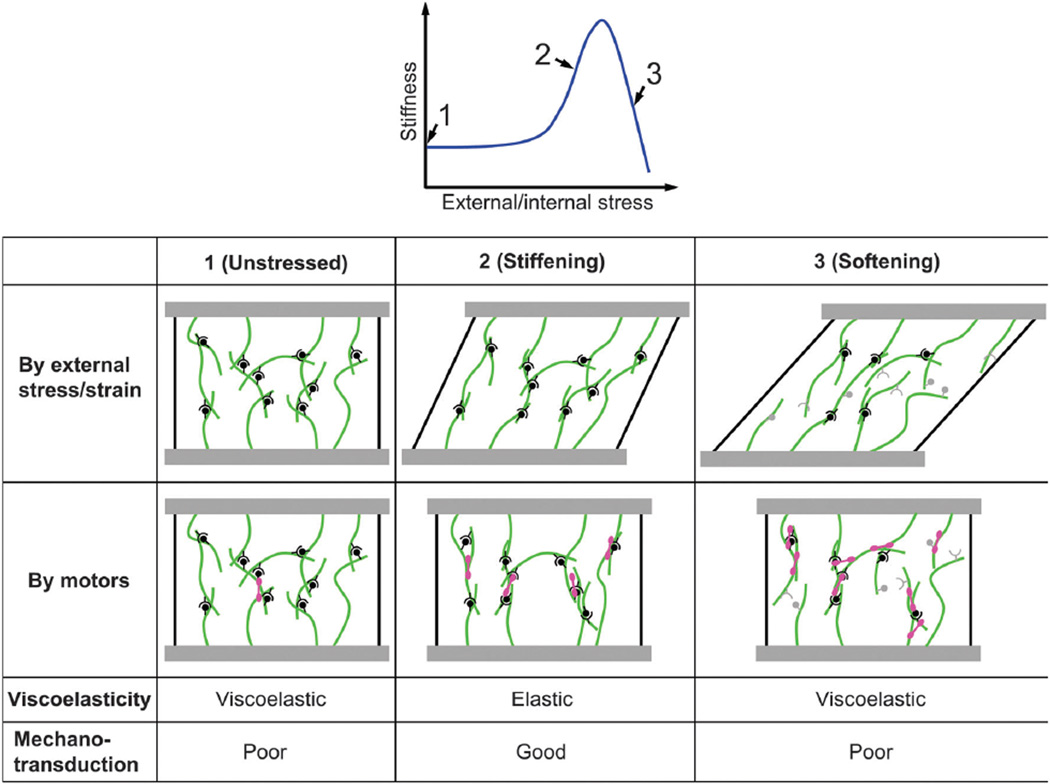
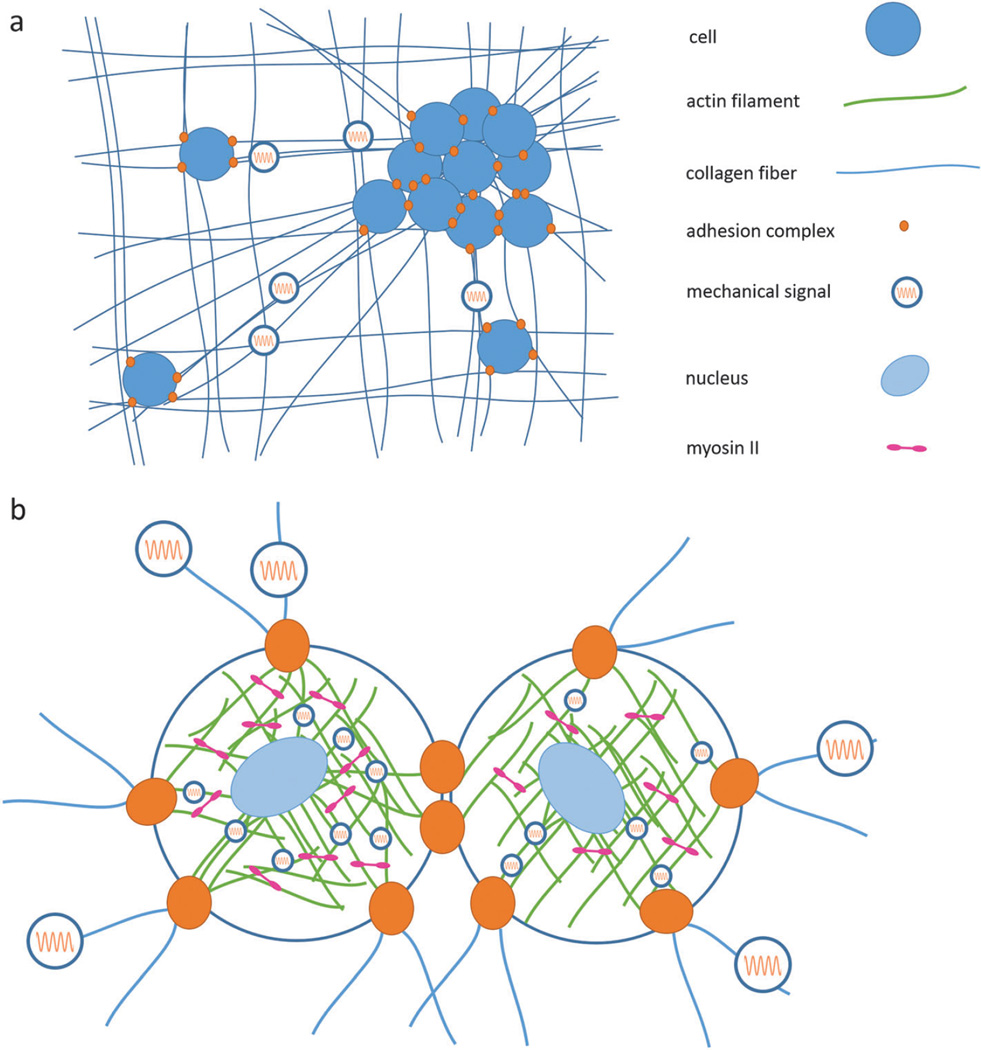
References
-
- Szabó B, Szöllösi GJ, Gönci B, Jurányi Z, Selmeczi D, Vicsek T. Phase transition in the collective migration of tissue cells: experiment and model, Phys. Rev. E: Stat., Nonlinear. Soft Matter Phys. 2006;74:061908. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
