

PDGFRα signaling drives adipose tissue fibrosis by targeting progenitor cell plasticity
- PMID: 26019175
- PMCID: PMC4470280
- DOI: 10.1101/gad.260554.115
PDGFRα signaling drives adipose tissue fibrosis by targeting progenitor cell plasticity
Abstract
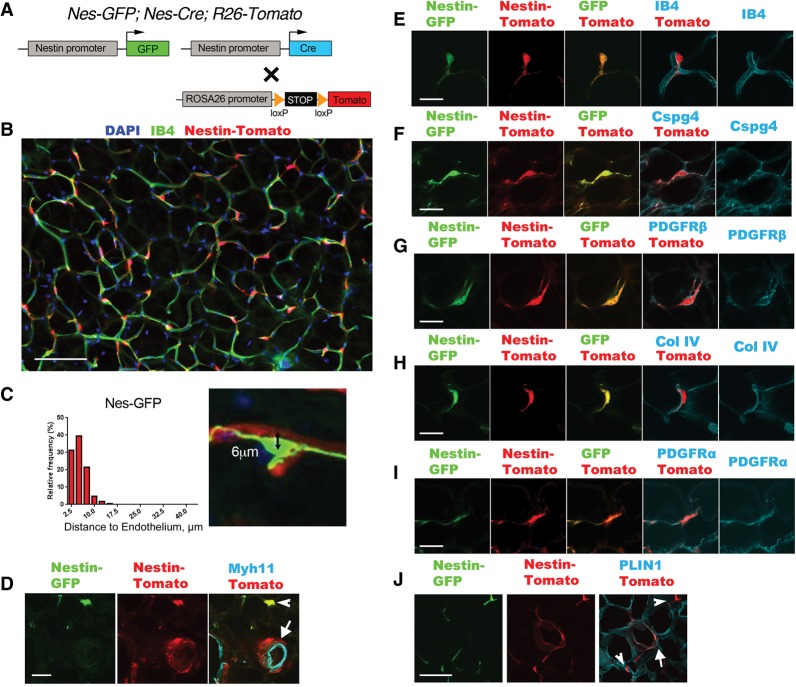
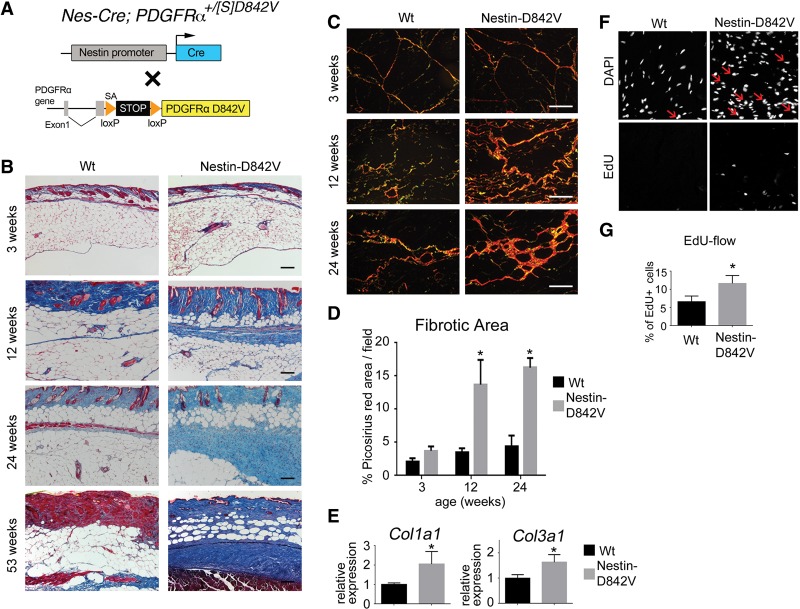
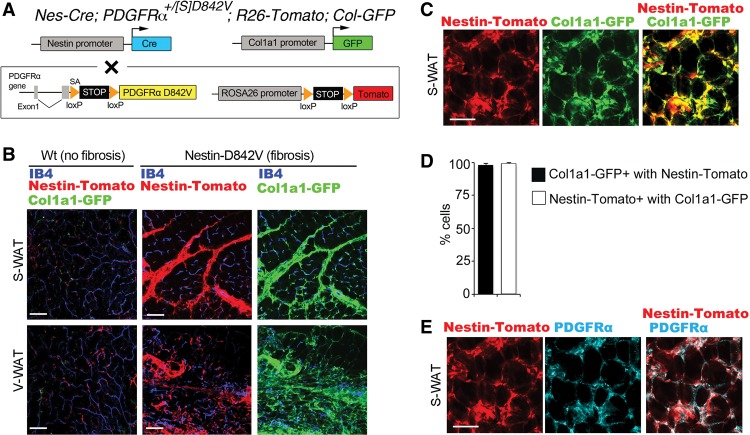
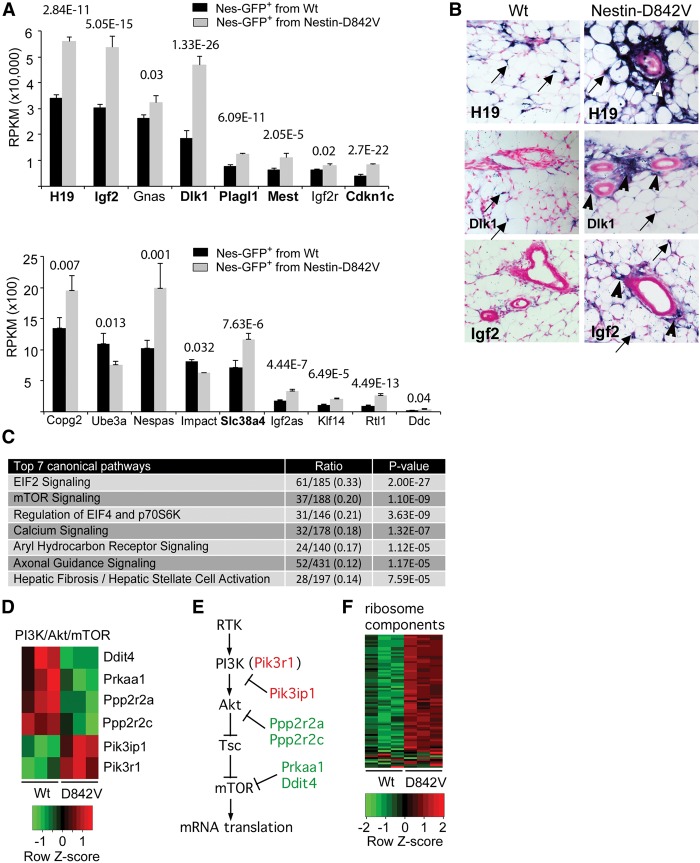
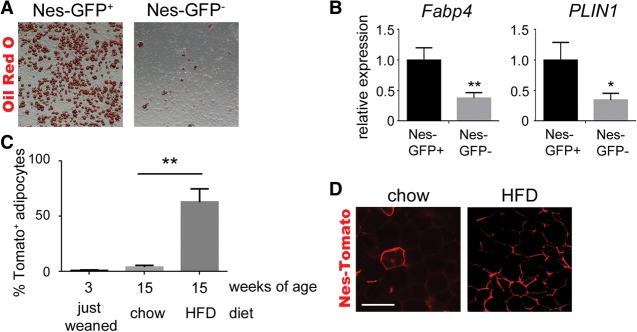
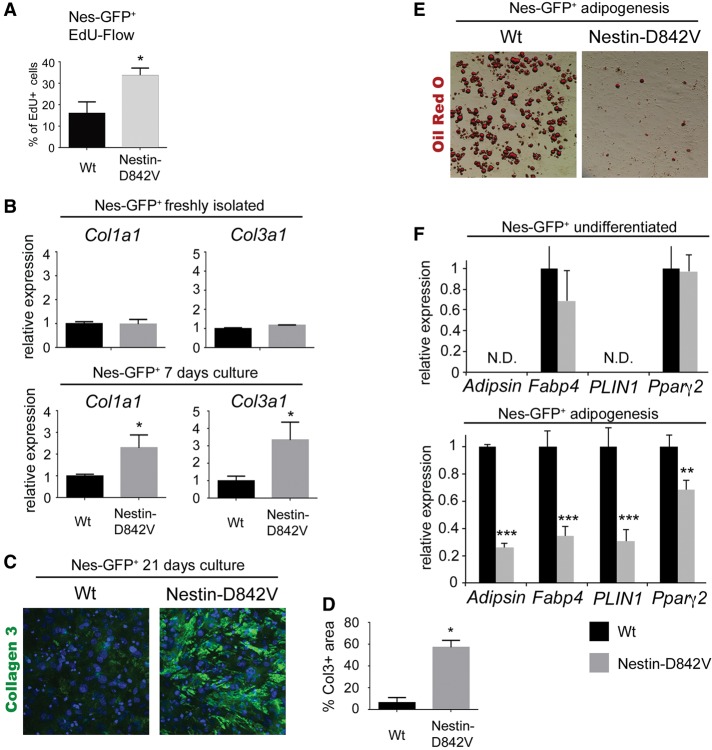
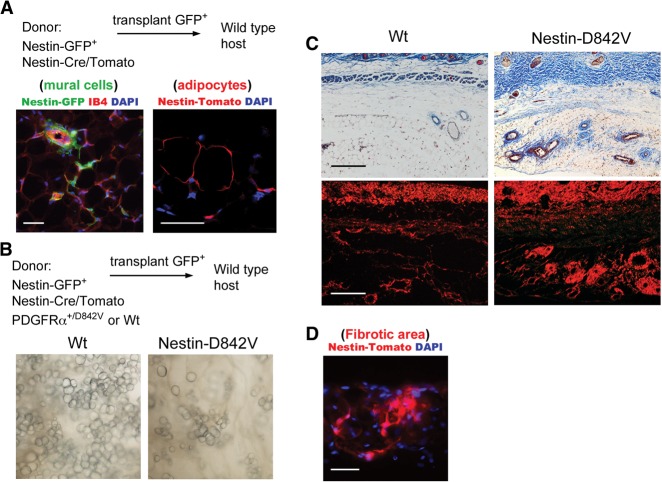
Fibrosis is a common disease process in which profibrotic cells disturb organ function by secreting disorganized extracellular matrix (ECM). Adipose tissue fibrosis occurs during obesity and is associated with metabolic dysfunction, but how profibrotic cells originate is still being elucidated. Here, we use a developmental model to investigate perivascular cells in white adipose tissue (WAT) and their potential to cause organ fibrosis. We show that a Nestin-Cre transgene targets perivascular cells (adventitial cells and pericyte-like cells) in WAT, and Nestin-GFP specifically labels pericyte-like cells. Activation of PDGFRα signaling in perivascular cells causes them to transition into ECM-synthesizing profibrotic cells. Before this transition occurs, PDGFRα signaling up-regulates mTOR signaling and ribosome biogenesis pathways and perturbs the expression of a network of epigenetically imprinted genes that have been implicated in cell growth and tissue homeostasis. Isolated Nestin-GFP(+) cells differentiate into adipocytes ex vivo and form WAT when transplanted into recipient mice. However, PDGFRα signaling opposes adipogenesis and generates profibrotic cells instead, which leads to fibrotic WAT in transplant experiments. These results identify perivascular cells as fibro/adipogenic progenitors in WAT and show that PDGFRα targets progenitor cell plasticity as a profibrotic mechanism.
Keywords: Nestin; adipogenesis; fibrosis; imprinting; pericyte; platelet-derived growth factor.
© 2015 Iwayama et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Akhmetshina A, Venalis P, Dees C, Busch N, Zwerina J, Schett G, Distler O, Distler JH. 2009. Treatment with imatinib prevents fibrosis in different preclinical models of systemic sclerosis and induces regression of established fibrosis. Arthritis Rheum 60: 219–224. - PubMed
-
- Armulik A, Genove G, Betsholtz C. 2011. Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev Cell 21: 193–215. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous