

The Cullin 4A/B-DDB1-Cereblon E3 Ubiquitin Ligase Complex Mediates the Degradation of CLC-1 Chloride Channels
- PMID: 26021757
- PMCID: PMC4448132
- DOI: 10.1038/srep10667
The Cullin 4A/B-DDB1-Cereblon E3 Ubiquitin Ligase Complex Mediates the Degradation of CLC-1 Chloride Channels
Abstract
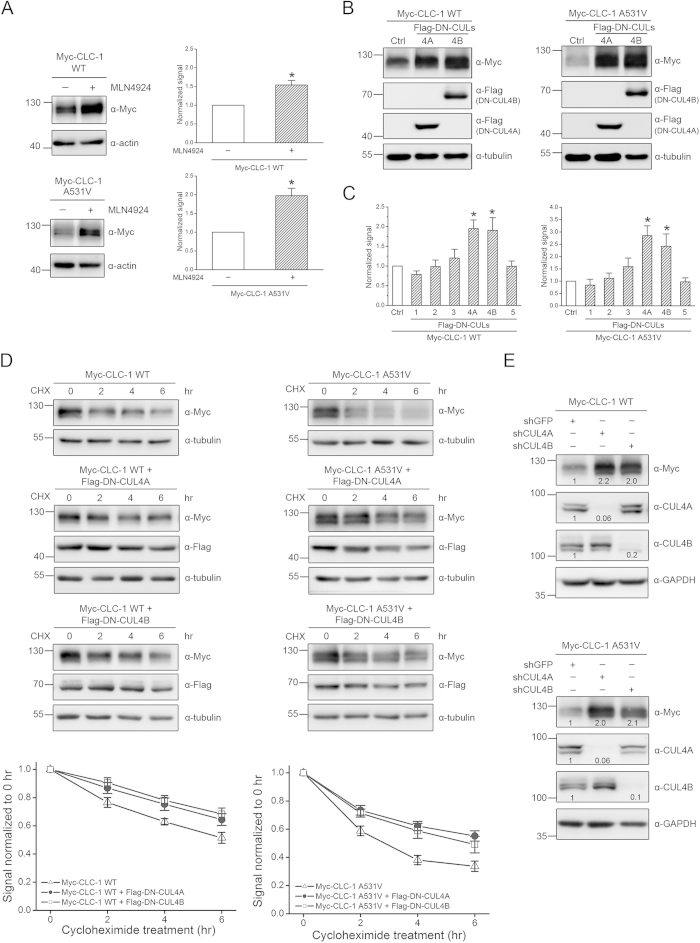
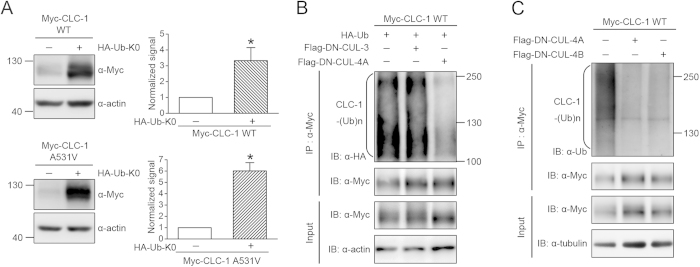
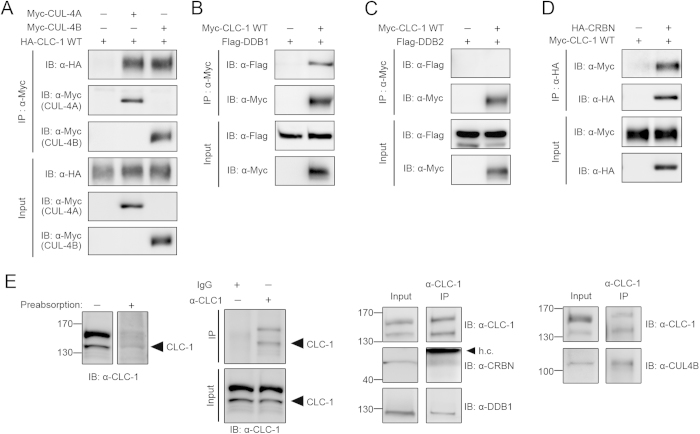
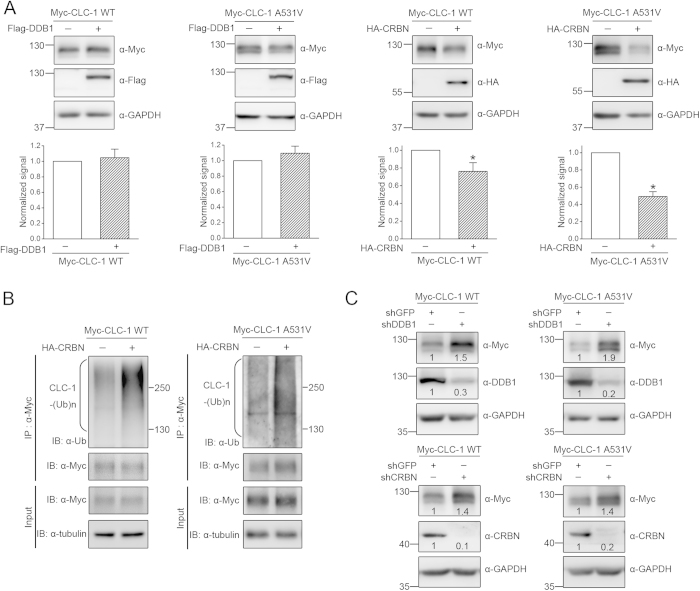
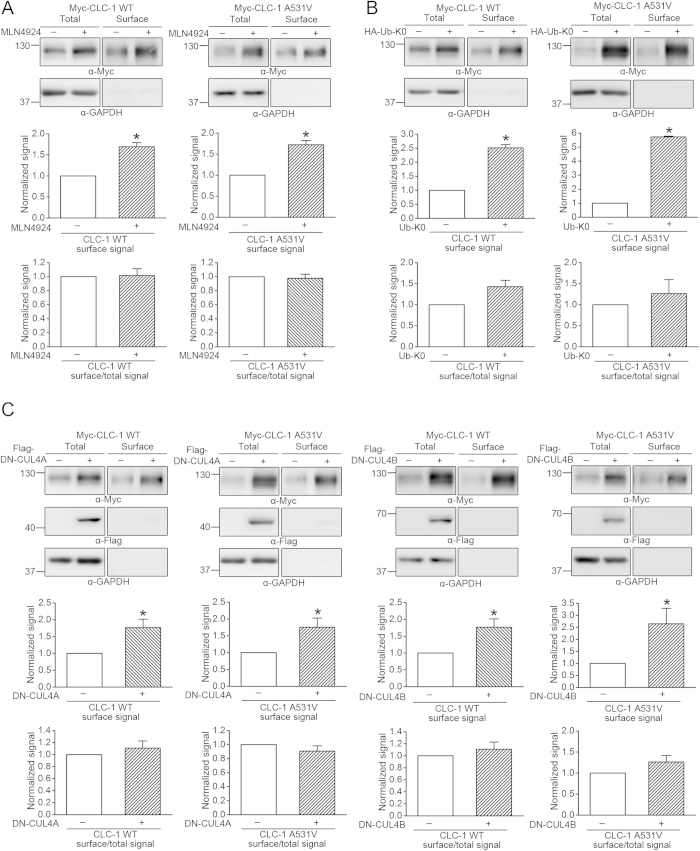
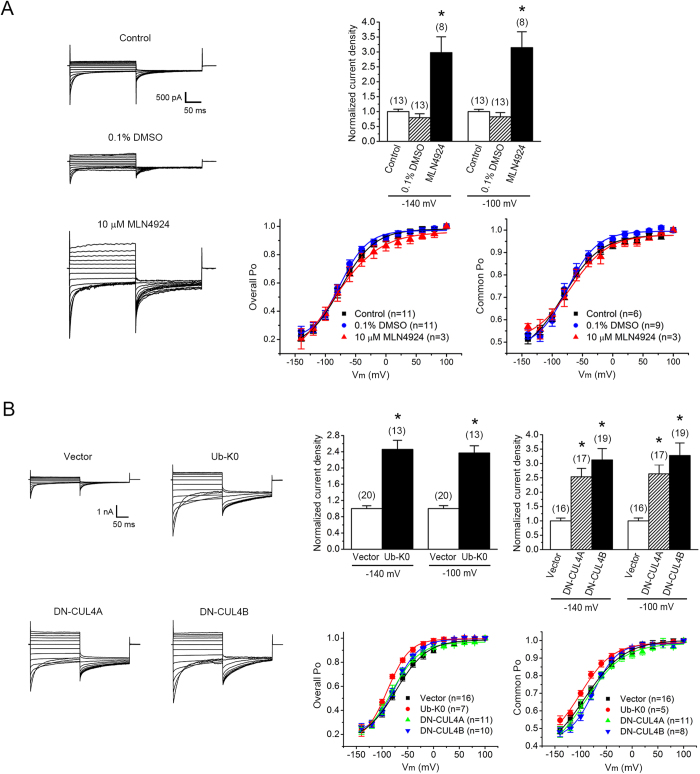
Voltage-gated CLC-1 chloride channels play a critical role in controlling the membrane excitability of skeletal muscles. Mutations in human CLC-1 channels have been linked to the hereditary muscle disorder myotonia congenita. We have previously demonstrated that disease-associated CLC-1 A531V mutant protein may fail to pass the endoplasmic reticulum quality control system and display enhanced protein degradation as well as defective membrane trafficking. Currently the molecular basis of protein degradation for CLC-1 channels is virtually unknown. Here we aim to identify the E3 ubiquitin ligase of CLC-1 channels. The protein abundance of CLC-1 was notably enhanced in the presence of MLN4924, a specific inhibitor of cullin-RING E3 ligases. Subsequent investigation with dominant-negative constructs against specific subtypes of cullin-RING E3 ligases suggested that CLC-1 seemed to serve as the substrate for cullin 4A (CUL4A) and 4B (CUL4B). Biochemical examinations further indicated that CUL4A/B, damage-specific DNA binding protein 1 (DDB1), and cereblon (CRBN) appeared to co-exist in the same protein complex with CLC-1. Moreover, suppression of CUL4A/B E3 ligase activity significantly enhanced the functional expression of the A531V mutant. Our data are consistent with the idea that the CUL4A/B-DDB1-CRBN complex catalyses the polyubiquitination and thus controls the degradation of CLC-1 channels.
Figures
References
-
- Bretag A. H. Muscle chloride channels. Physiol Rev 67, 618–724 (1987). - PubMed
-
- Dulhunty A. F. Distribution of potassium and chloride permeability over the surface and T-tubule membranes of mammalian skeletal muscle. J Membr Biol 45, 293–310 (1979). - PubMed
-
- Jurkat-Rott K., Lerche H. & Lehmann-Horn F. Skeletal muscle channelopathies. J Neurol 249, 1493–1502 (2002). - PubMed
-
- Lossin C. & George A. L. Jr. Myotonia congenita. Adv Genet 63, 25–55 (2008). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
