

Dnmt1 is essential to maintain progenitors in the perinatal intestinal epithelium
- PMID: 26023099
- PMCID: PMC4483766
- DOI: 10.1242/dev.117341
Dnmt1 is essential to maintain progenitors in the perinatal intestinal epithelium
Abstract
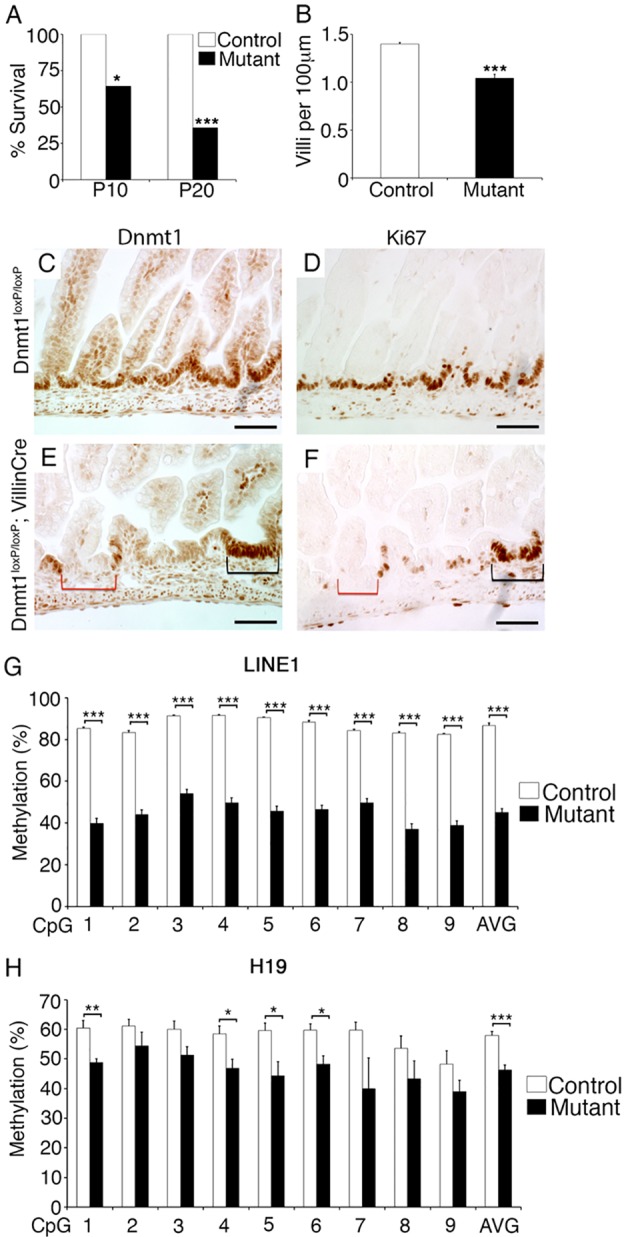
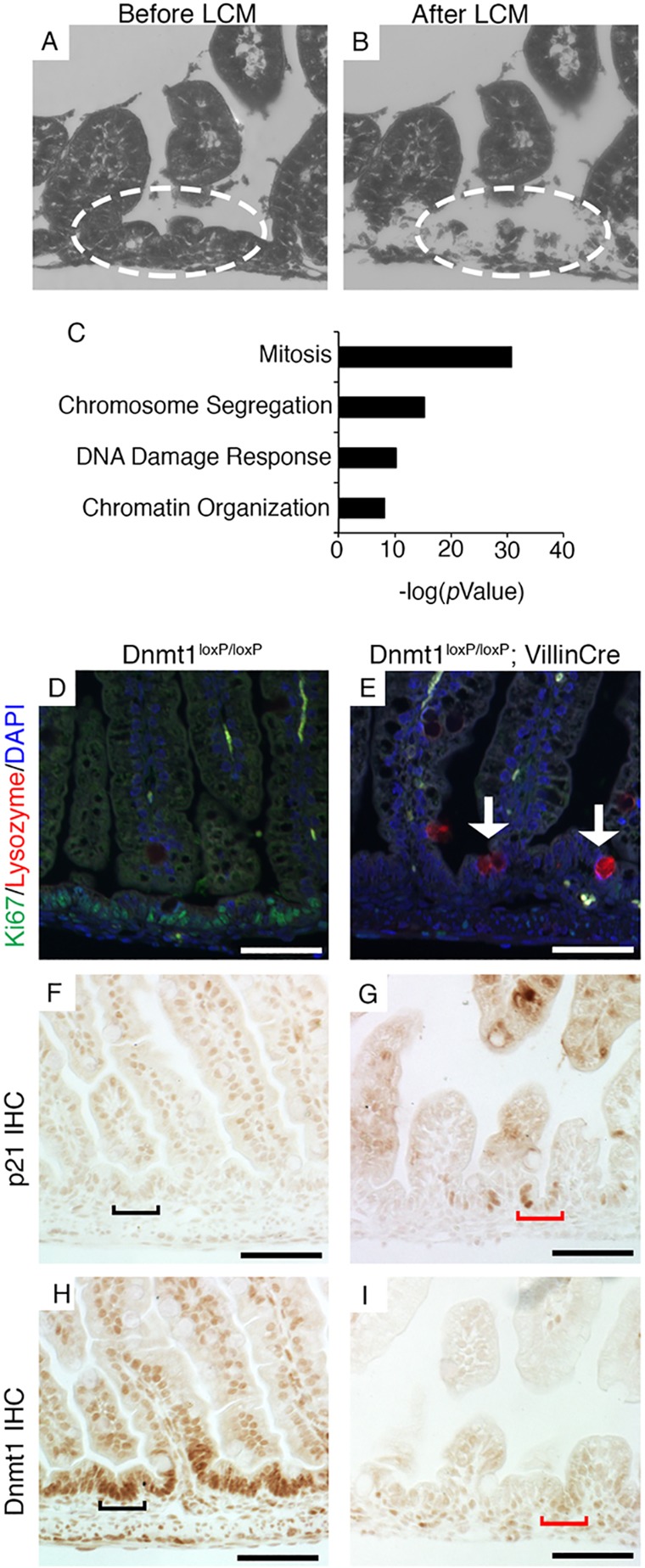
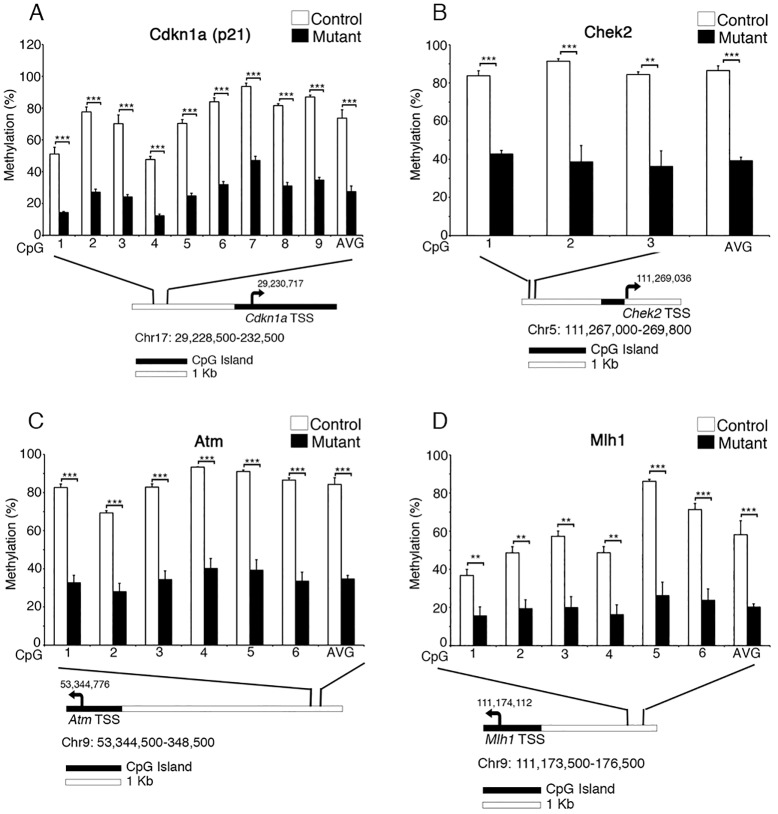
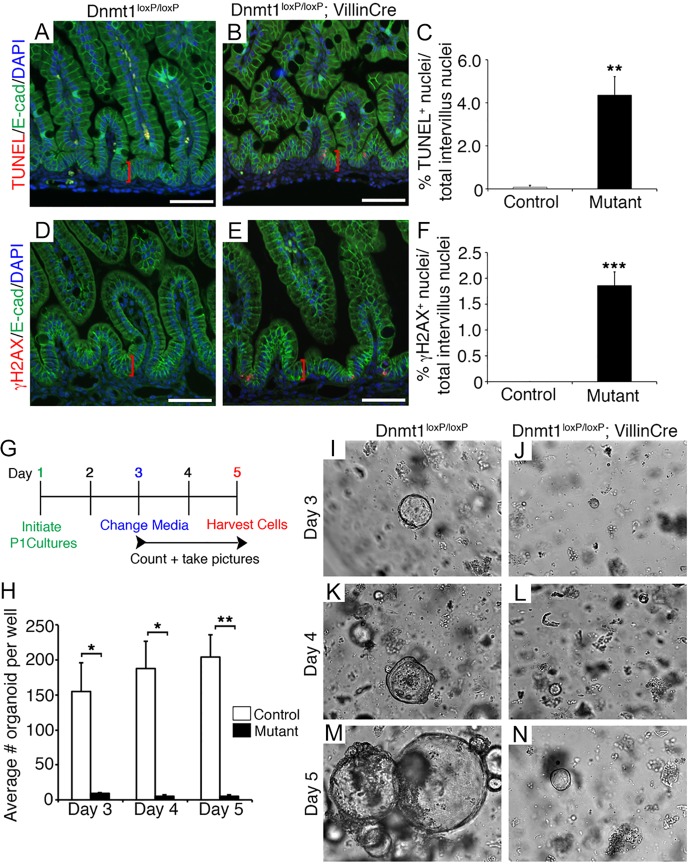
The DNA methyltransferase Dnmt1 maintains DNA methylation patterns and genomic stability in several in vitro cell systems. Ablation of Dnmt1 in mouse embryos causes death at the post-gastrulation stage; however, the functions of Dnmt1 and DNA methylation in organogenesis remain unclear. Here, we report that Dnmt1 is crucial during perinatal intestinal development. Loss of Dnmt1 in intervillus progenitor cells causes global hypomethylation, DNA damage, premature differentiation, apoptosis and, consequently, loss of nascent villi. We further confirm the crucial role of Dnmt1 during crypt development using the in vitro organoid culture system, and illustrate a clear differential requirement for Dnmt1 in immature versus mature organoids. These results demonstrate an essential role for Dnmt1 in maintaining genomic stability during intestinal development and the establishment of intestinal crypts.
Keywords: DNA damage response; DNA methylation; Intestinal epithelial development.
© 2015. Published by The Company of Biologists Ltd.
Figures
References
-
- Ahuja N., Mohan A. L., Li Q., Stolker J. M., Herman J. G., Hamilton S. R., Baylin S. B. and Issa J. P. (1997). Association between CpG island methylation and microsatellite instability in colorectal cancer. Cancer Res. 57, 3370-3374. - PubMed
-
- Allan L. A., Duhig T., Read M. and Fried M. (2000). The p21(WAF1/CIP1) promoter is methylated in Rat-1 cells: stable restoration of p53-dependent p21(WAF1/CIP1) expression after transfection of a genomic clone containing the p21(WAF1/CIP1) gene. Mol. Cell. Biol. 20, 1291-1298. 10.1128/MCB.20.4.1291-1298.2000 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
