

The Fifth Transmembrane Segment of Cystic Fibrosis Transmembrane Conductance Regulator Contributes to Its Anion Permeation Pathway
- PMID: 26024338
- PMCID: PMC6510985
- DOI: 10.1021/acs.biochem.5b00427
The Fifth Transmembrane Segment of Cystic Fibrosis Transmembrane Conductance Regulator Contributes to Its Anion Permeation Pathway
Abstract
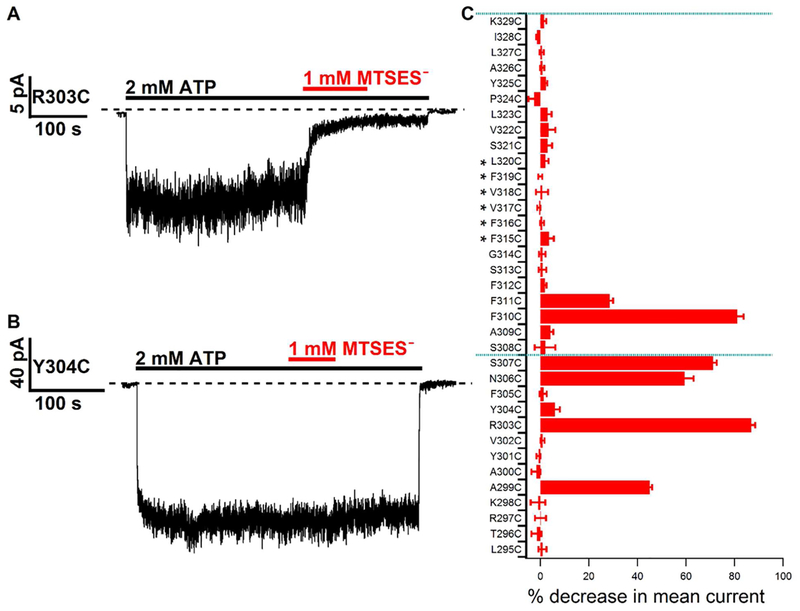
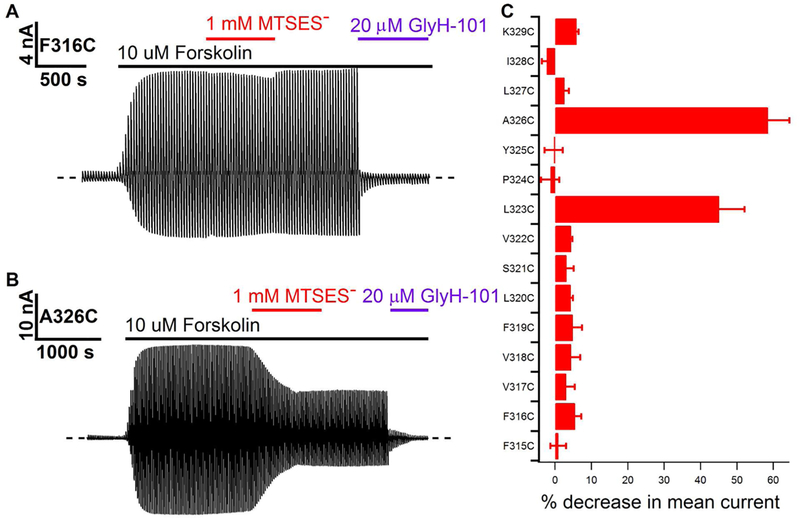
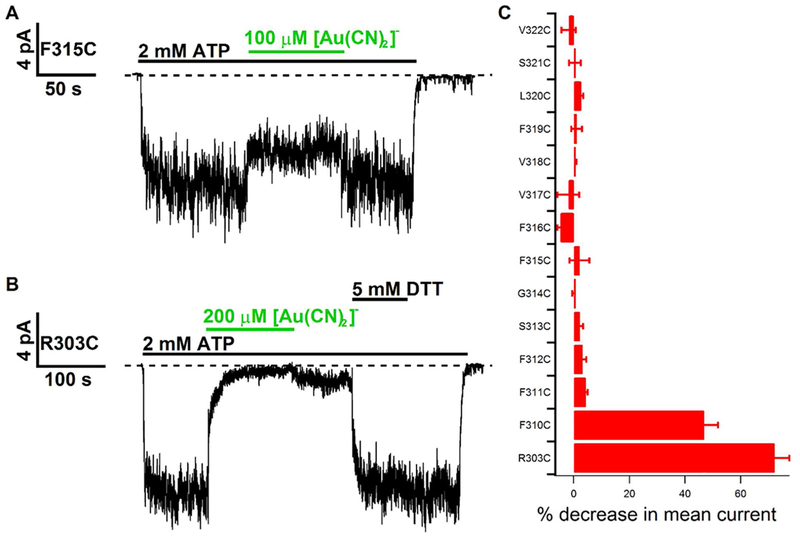
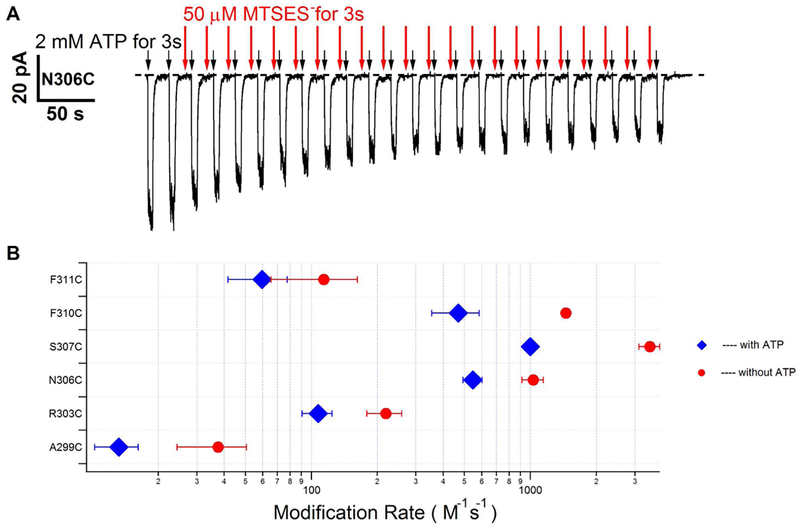
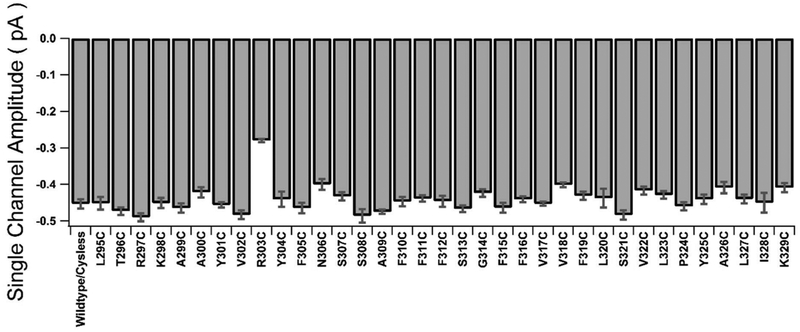
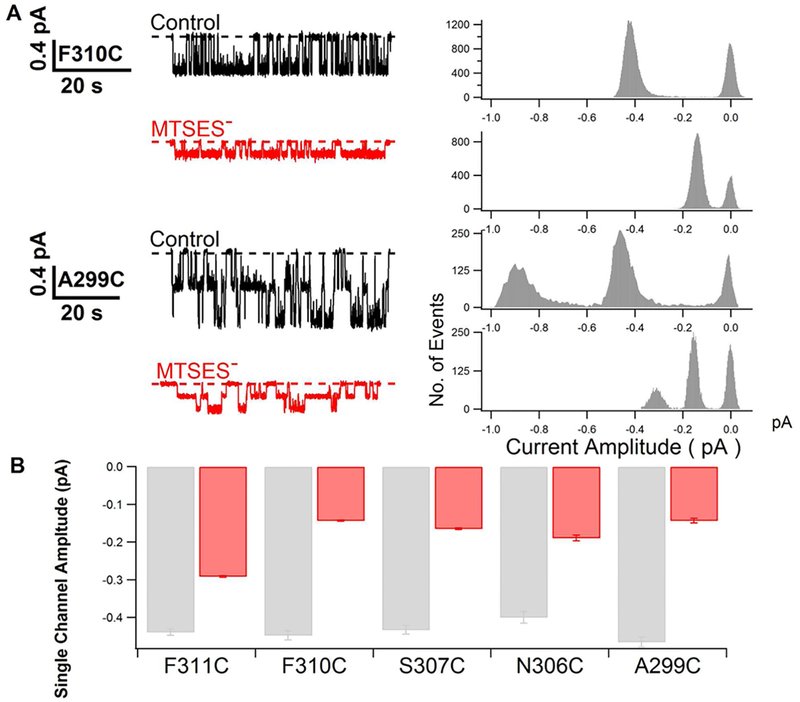
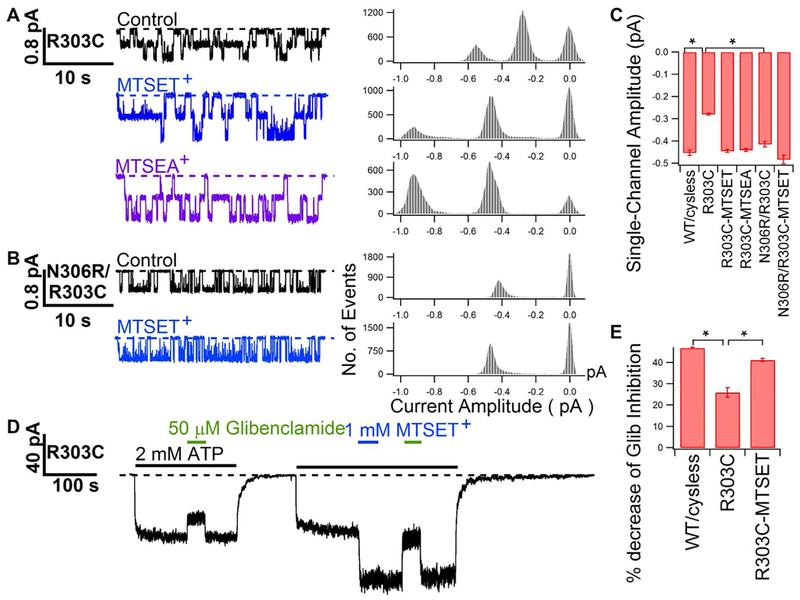
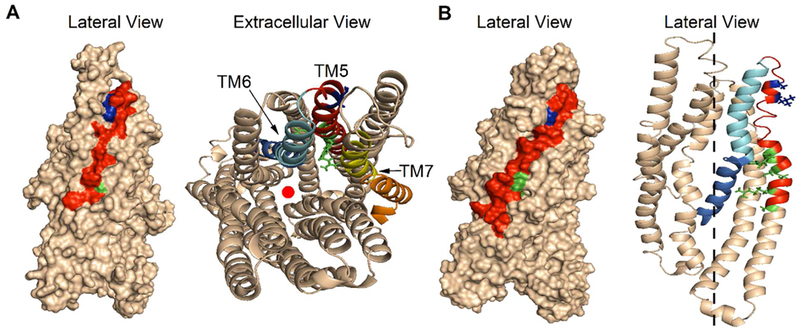
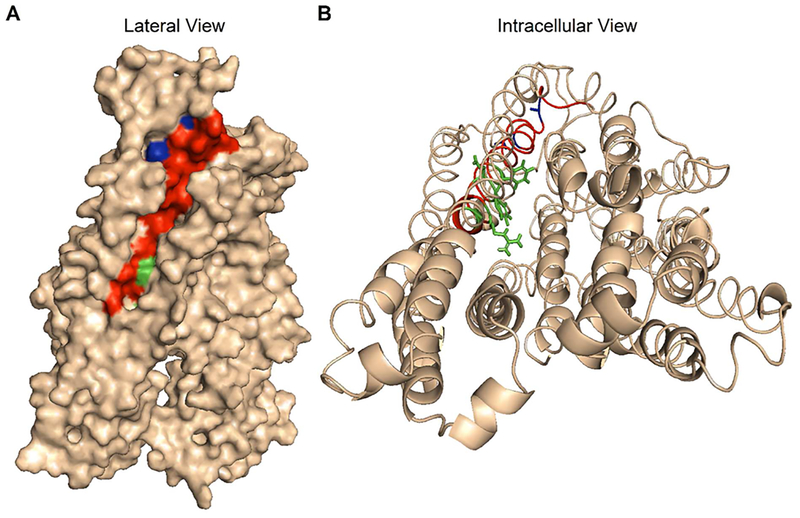
Previous studies have identified several transmembrane segments (TMs), including TM1, TM3, TM6, TM9, TM11, and TM12, as pore-lining segments in cystic fibrosis transmembrane conductance regulator (CFTR), but the role of TM5 in pore construction remains controversial. In this study, we employed substituted cysteine accessibility methodology (SCAM) to screen the entire TM5 defined by the original topology model and its cytoplasmic extension in a Cysless background. We found six positions (A299, R303, N306, S307, F310, and F311) where engineered cysteines react to intracellular 2-sulfonatoethyl methanethiosulfonate (MTSES⁻). Quantification of the modification rate of engineered cysteines in the presence or absence of ATP suggests that these six residues are accessible in both the open and closed states. Whole-cell experiments with external MTSES⁻ identified only two positive positions (L323 and A326), resulting in a segment containing 11 consecutive amino acids, where substituted cysteines respond to neither internal nor external MTSES⁻, a unique feature not seen previously in CFTR's pore-lining segments. The observation that these positions are inaccessible to channel-permeant thiol-specific reagent [Au(CN)₂]⁻ suggests that this segment of TM5 between F311 and L323 is concealed from the pore by other TMs and/or lipid bilayers. In addition, our data support the idea that the positively charged arginine at position 303 poses a pure electrostatic action in determining the single-channel current amplitude of CFTR and the effect of an open-channel blocker glibencalmide. Collectively, we conclude that the cytoplasmic portion of CFTR's TM5 lines the pore. Our functional data are remarkably consistent with predicted structural arrangements of TM5 in some homology models of CFTR.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
Cysteine scanning of CFTR's first transmembrane segment reveals its plausible roles in gating and permeation.Biophys J. 2013 Feb 19;104(4):786-97. doi: 10.1016/j.bpj.2012.12.048. Biophys J. 2013. PMID: 23442957 Free PMC article.
-
Changes in accessibility of cytoplasmic substances to the pore associated with activation of the cystic fibrosis transmembrane conductance regulator chloride channel.J Biol Chem. 2010 Oct 15;285(42):32126-40. doi: 10.1074/jbc.M110.113332. Epub 2010 Jul 30. J Biol Chem. 2010. PMID: 20675380 Free PMC article.
-
Spatial positioning of CFTR's pore-lining residues affirms an asymmetrical contribution of transmembrane segments to the anion permeation pathway.J Gen Physiol. 2016 May;147(5):407-22. doi: 10.1085/jgp.201511557. J Gen Physiol. 2016. PMID: 27114613 Free PMC article.
-
CFTR: what's it like inside the pore?J Exp Zool A Comp Exp Biol. 2003 Nov 1;300(1):69-75. doi: 10.1002/jez.a.10311. J Exp Zool A Comp Exp Biol. 2003. PMID: 14598388 Review.
-
Characterizing diverse orthologues of the cystic fibrosis transmembrane conductance regulator protein for structural studies.Biochem Soc Trans. 2015 Oct;43(5):894-900. doi: 10.1042/BST20150081. Biochem Soc Trans. 2015. PMID: 26517900 Review.
Cited by
-
Functional organization of cytoplasmic portals controlling access to the cystic fibrosis transmembrane conductance regulator (CFTR) chloride channel pore.J Biol Chem. 2018 Apr 13;293(15):5649-5658. doi: 10.1074/jbc.RA117.001373. Epub 2018 Feb 23. J Biol Chem. 2018. PMID: 29475947 Free PMC article.
-
Contribution of the eighth transmembrane segment to the function of the CFTR chloride channel pore.Cell Mol Life Sci. 2019 Jun;76(12):2411-2423. doi: 10.1007/s00018-019-03043-2. Epub 2019 Feb 13. Cell Mol Life Sci. 2019. PMID: 30758641 Free PMC article.
-
The molecular evolution of function in the CFTR chloride channel.J Gen Physiol. 2021 Dec 6;153(12):e202012625. doi: 10.1085/jgp.202012625. Epub 2021 Oct 14. J Gen Physiol. 2021. PMID: 34647973 Free PMC article.
-
Functional characterization reveals that zebrafish CFTR prefers to occupy closed channel conformations.PLoS One. 2018 Dec 31;13(12):e0209862. doi: 10.1371/journal.pone.0209862. eCollection 2018. PLoS One. 2018. PMID: 30596737 Free PMC article.
-
CFTR Modulators: From Mechanism to Targeted Therapeutics.Handb Exp Pharmacol. 2024;283:219-247. doi: 10.1007/164_2022_597. Handb Exp Pharmacol. 2024. PMID: 35972584
References
-
- Dean M, and Annilo T (2005) Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates. Annu. Rev. Genomics Hum. Genet 6, 123–142. - PubMed
-
- Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou JL, et al. (1989) Identification of the cystic fibrosis gene: Cloning and characterization of complementary DNA. Science 245, 1066–1073. - PubMed
-
- Bear CE, Li CH, Kartner N, Bridges RJ, Jensen TJ, Ramjeesingh M, and Riordan JR (1992) Purification and functional reconstitution of the cystic fibrosis transmembrane conductance regulator (CFTR). Cell 68, 809–818. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
