

Distinct circuit-dependent functions of presynaptic neurexin-3 at GABAergic and glutamatergic synapses
- PMID: 26030848
- PMCID: PMC4482778
- DOI: 10.1038/nn.4037
Distinct circuit-dependent functions of presynaptic neurexin-3 at GABAergic and glutamatergic synapses
Abstract
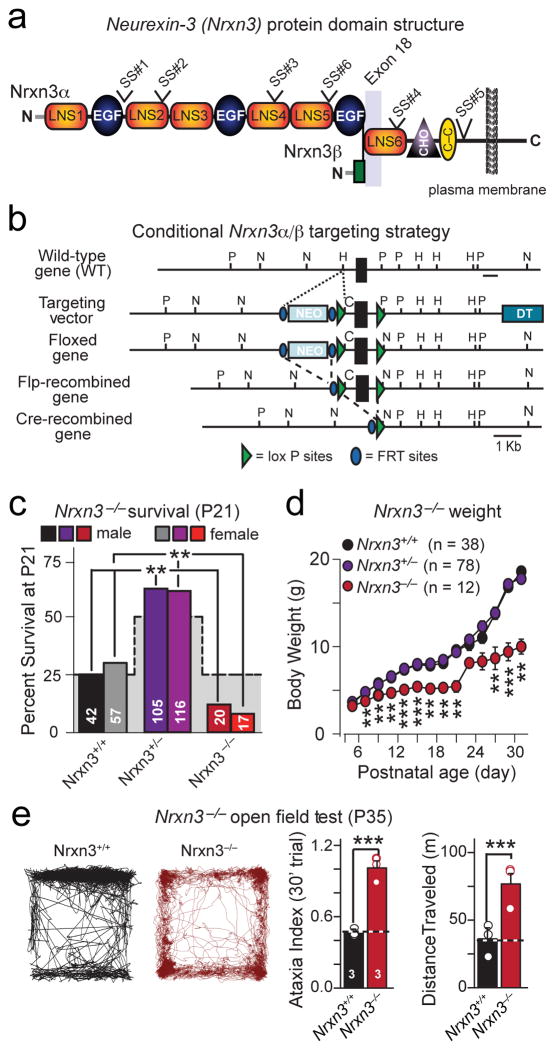
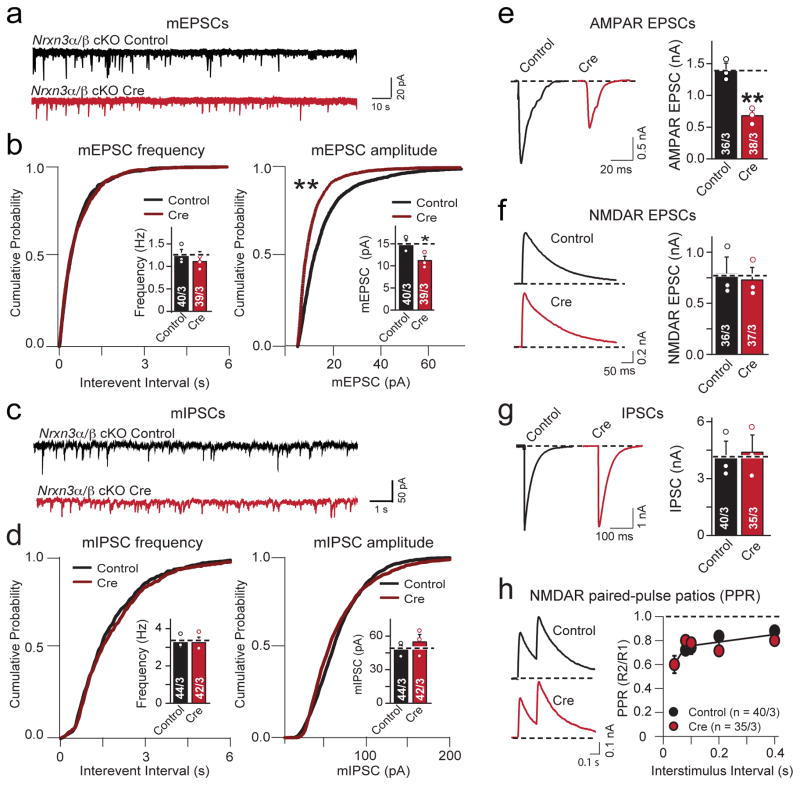
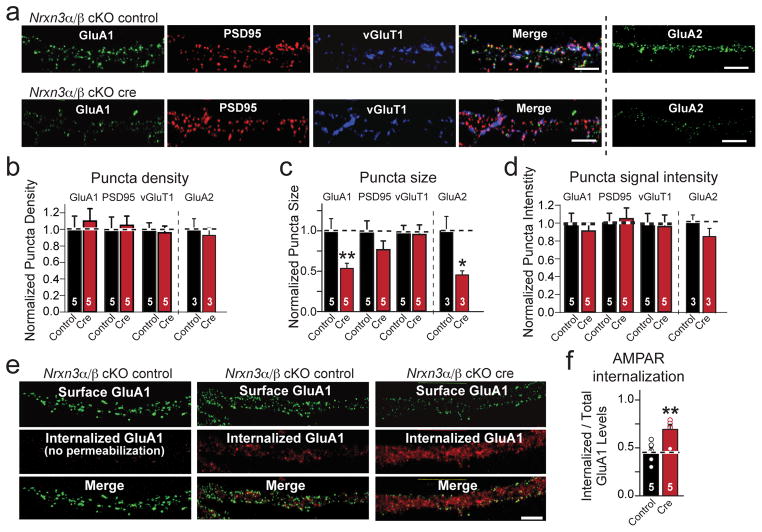
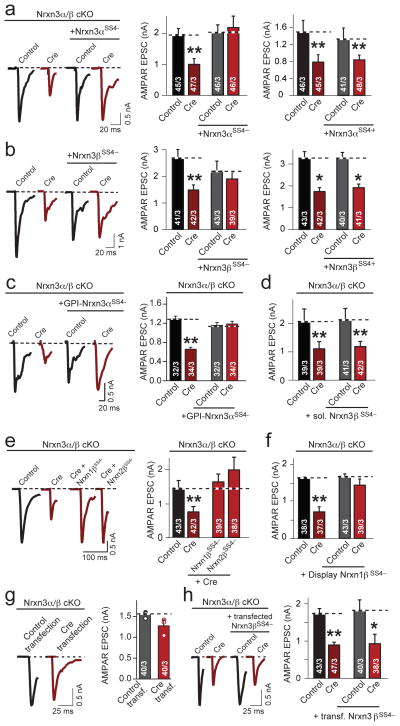
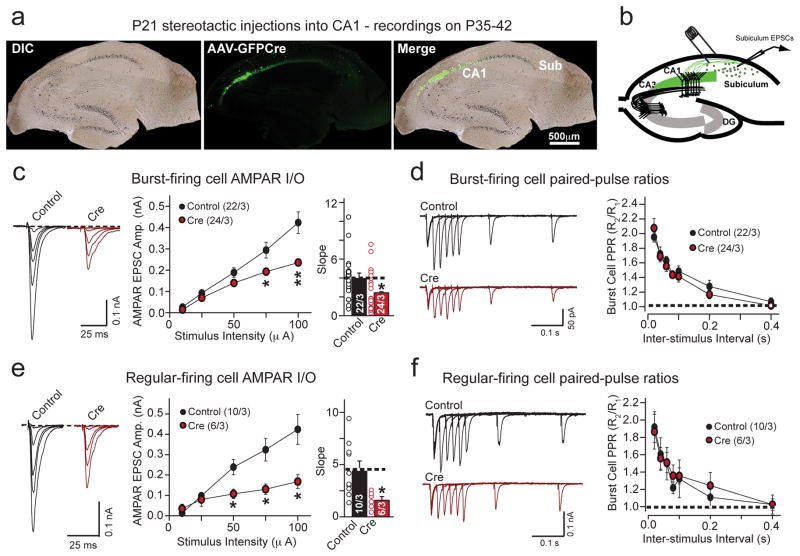
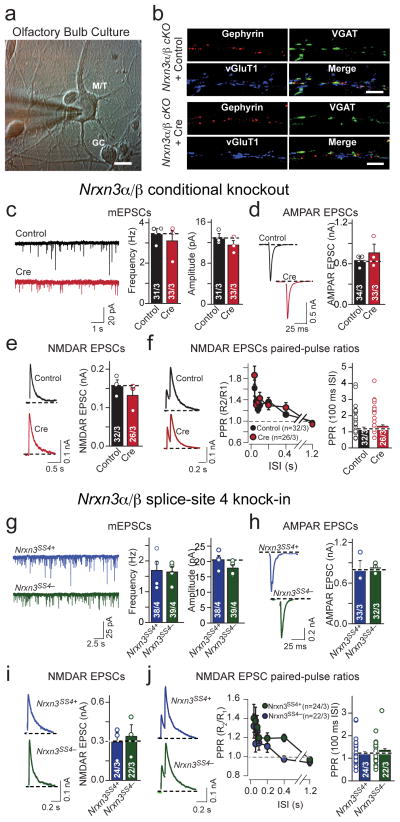
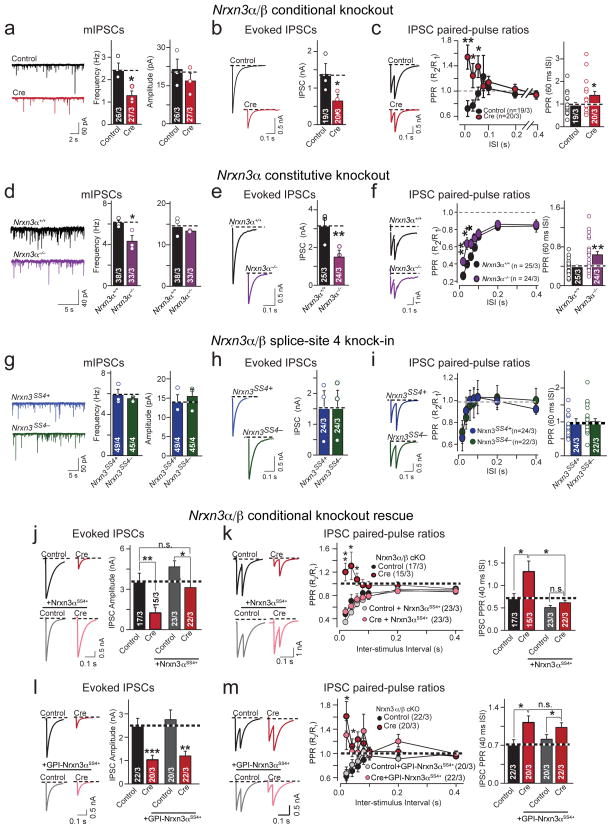
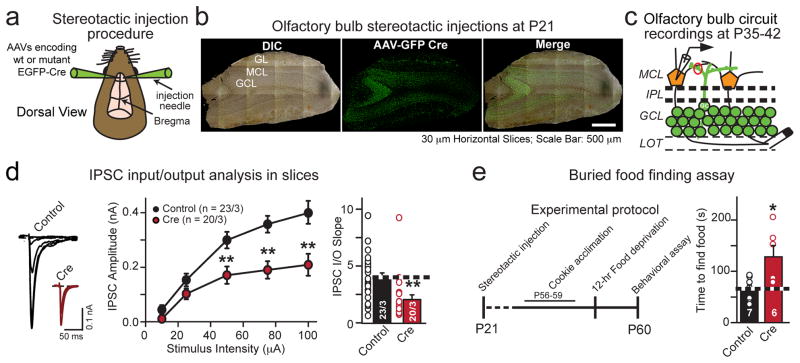
α- and β-neurexins are presynaptic cell-adhesion molecules whose general importance for synaptic transmission is well documented. The specific functions of neurexins, however, remain largely unknown because no conditional neurexin knockouts are available and targeting all α- and β-neurexins produced by a particular gene is challenging. Using newly generated constitutive and conditional knockout mice that target all neurexin-3α and neurexin-3β isoforms, we found that neurexin-3 was differentially required for distinct synaptic functions in different brain regions. Specifically, we found that, in cultured neurons and acute slices of the hippocampus, extracellular sequences of presynaptic neurexin-3 mediated trans-synaptic regulation of postsynaptic AMPA receptors. In cultured neurons and acute slices of the olfactory bulb, however, intracellular sequences of presynaptic neurexin-3 were selectively required for GABA release. Thus, our data indicate that neurexin-3 performs distinct essential pre- or postsynaptic functions in different brain regions by distinct mechanisms.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
