

BCL-3 expression promotes colorectal tumorigenesis through activation of AKT signalling
- PMID: 26033966
- PMCID: PMC4941180
- DOI: 10.1136/gutjnl-2014-308270
BCL-3 expression promotes colorectal tumorigenesis through activation of AKT signalling
Abstract
Objective: Colorectal cancer remains the fourth most common cause of cancer-related mortality worldwide. Here we investigate the role of nuclear factor-κB (NF-κB) co-factor B-cell CLL/lymphoma 3 (BCL-3) in promoting colorectal tumour cell survival.
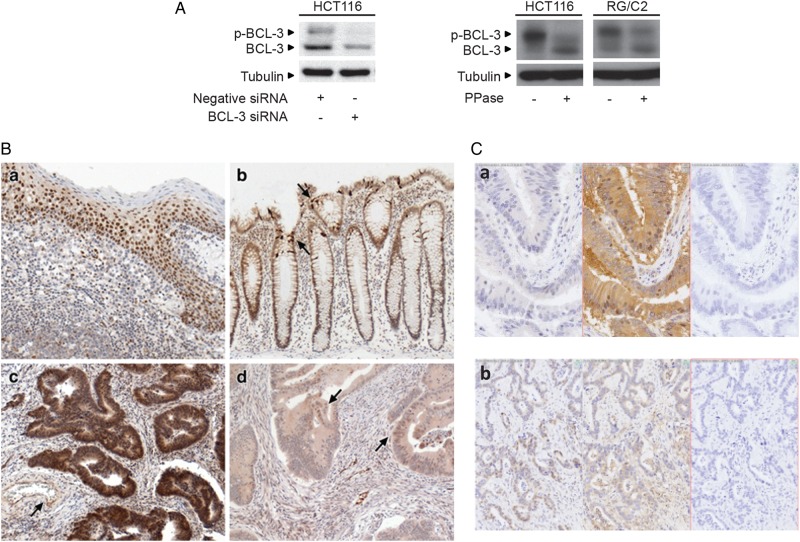
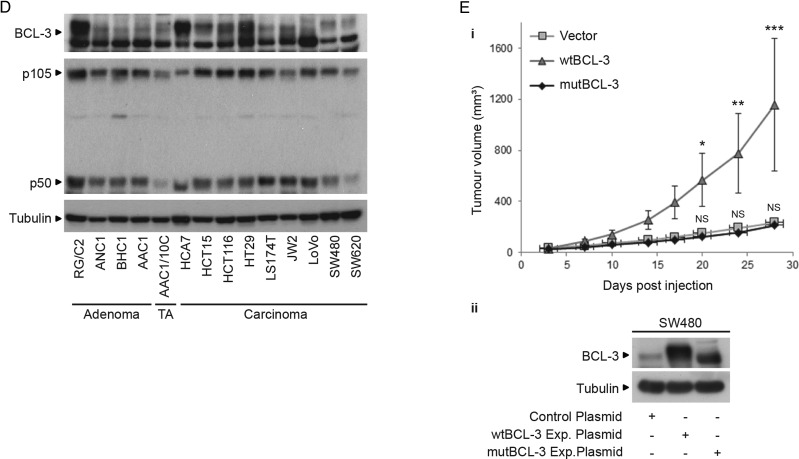
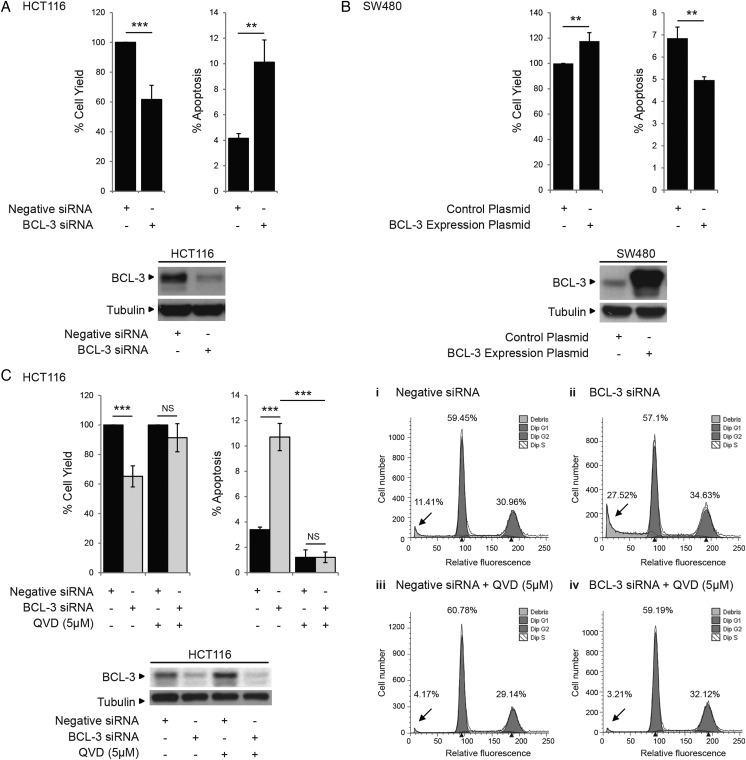
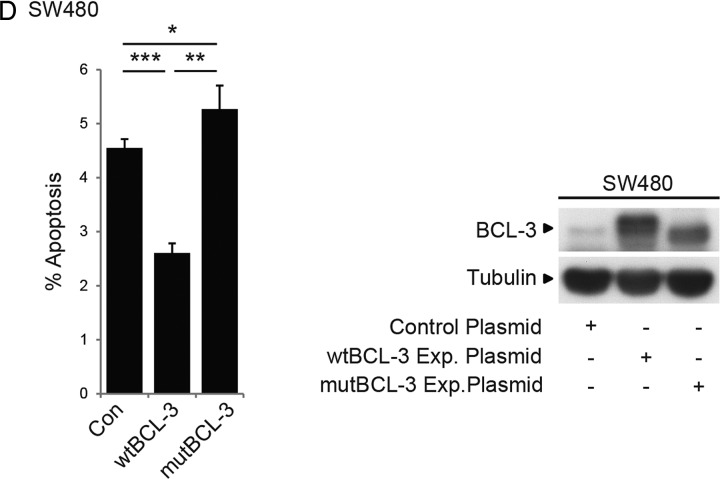
Design: Immunohistochemistry was carried out on 47 tumour samples and normal tissue from resection margins. The role of BCL-3/NF-κB complexes on cell growth was studied in vivo and in vitro using an siRNA approach and exogenous BCL-3 expression in colorectal adenoma and carcinoma cells. The question whether BCL-3 activated the AKT/protein kinase B (PKB) pathway in colorectal tumour cells was addressed by western blotting and confocal microscopy, and the ability of 5-aminosalicylic acid (5-ASA) to suppress BCL-3 expression was also investigated.
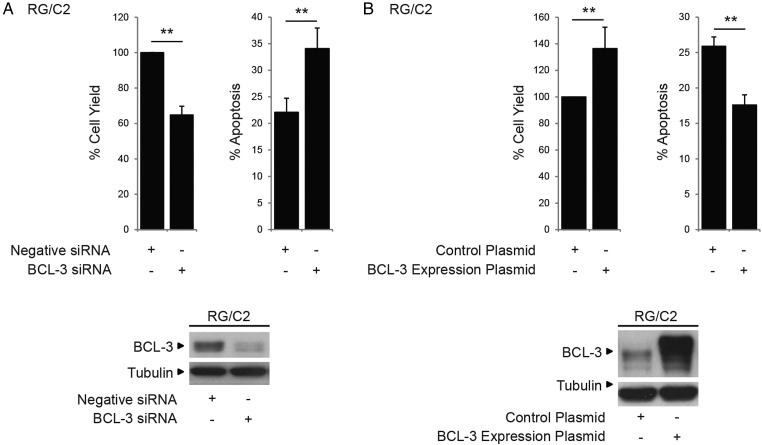
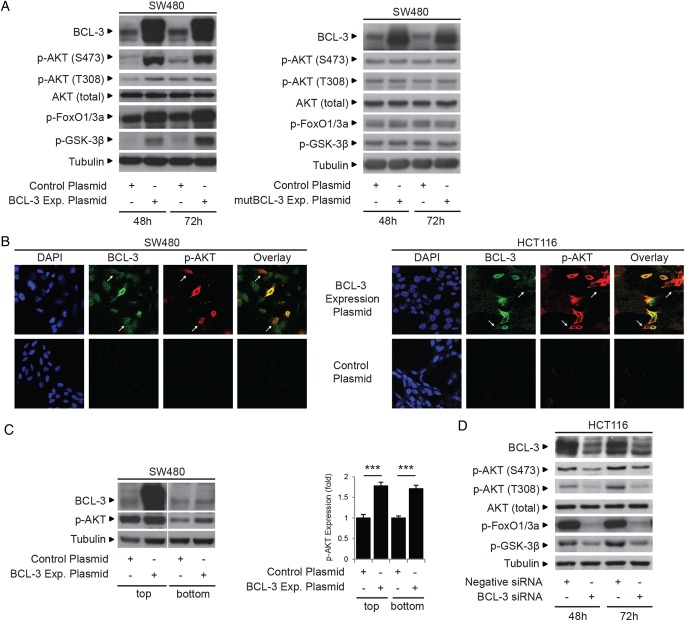
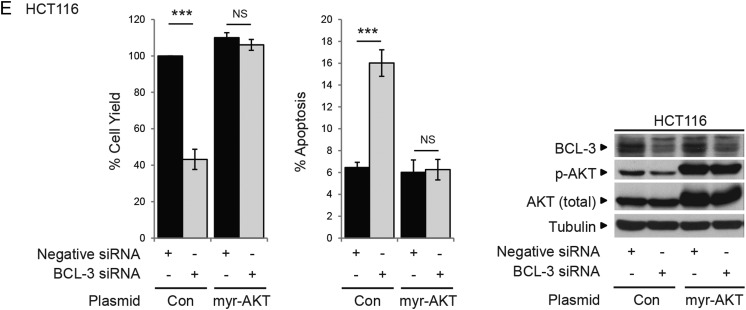
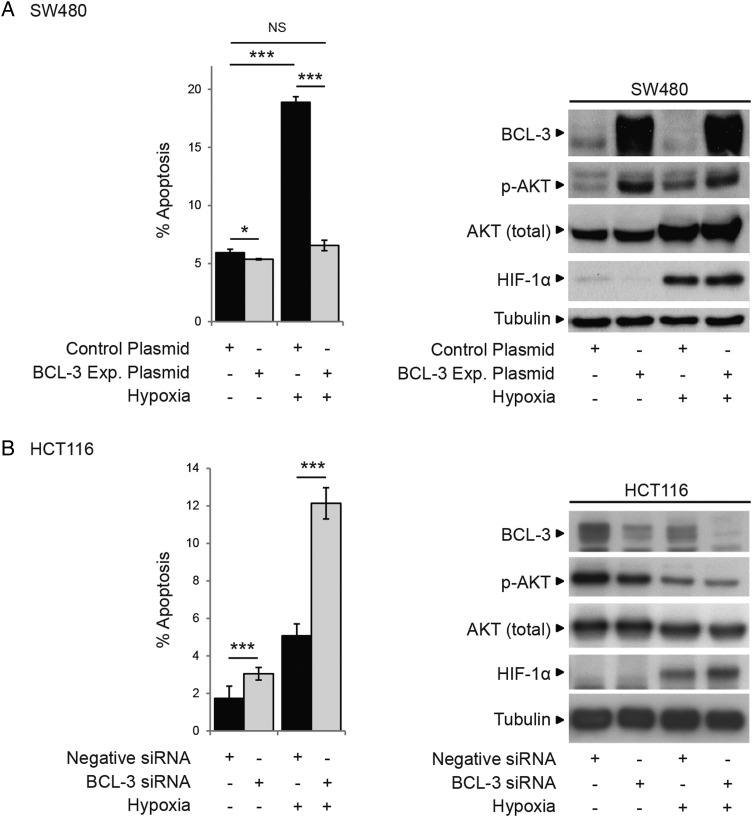
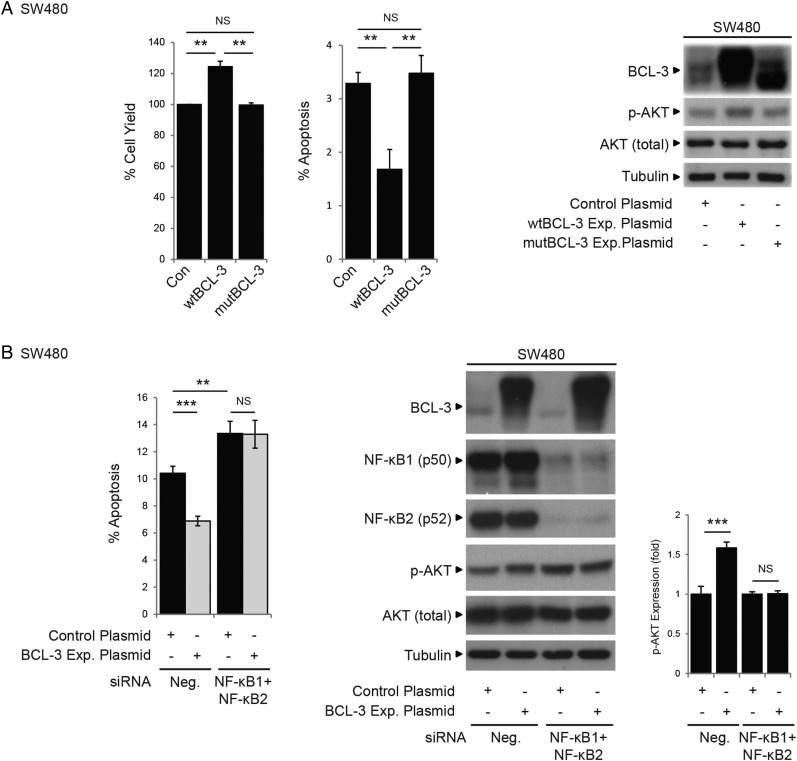
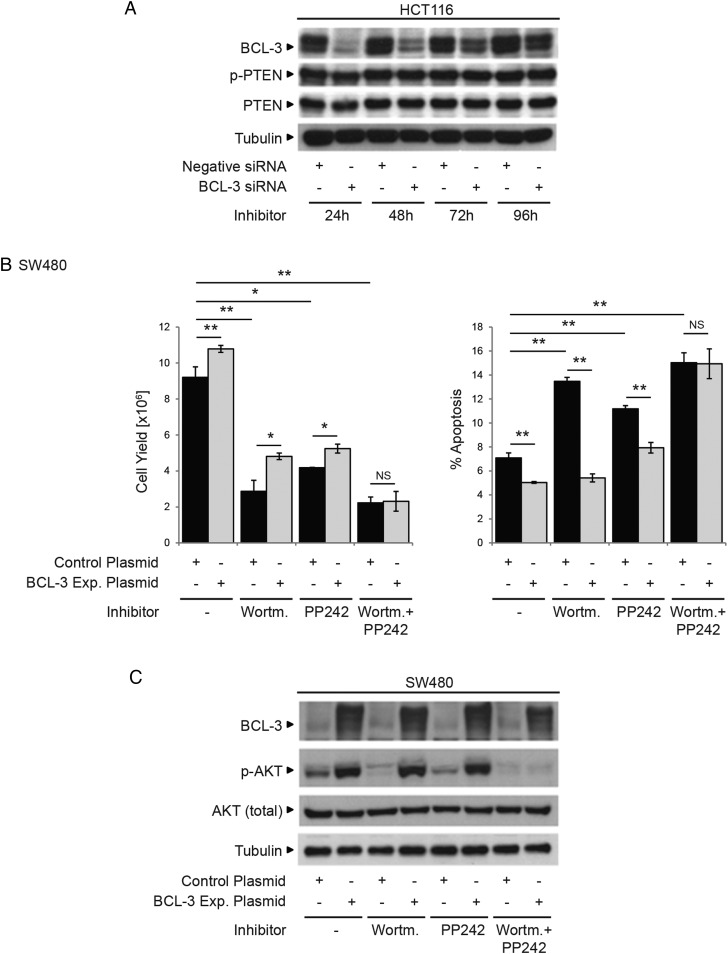
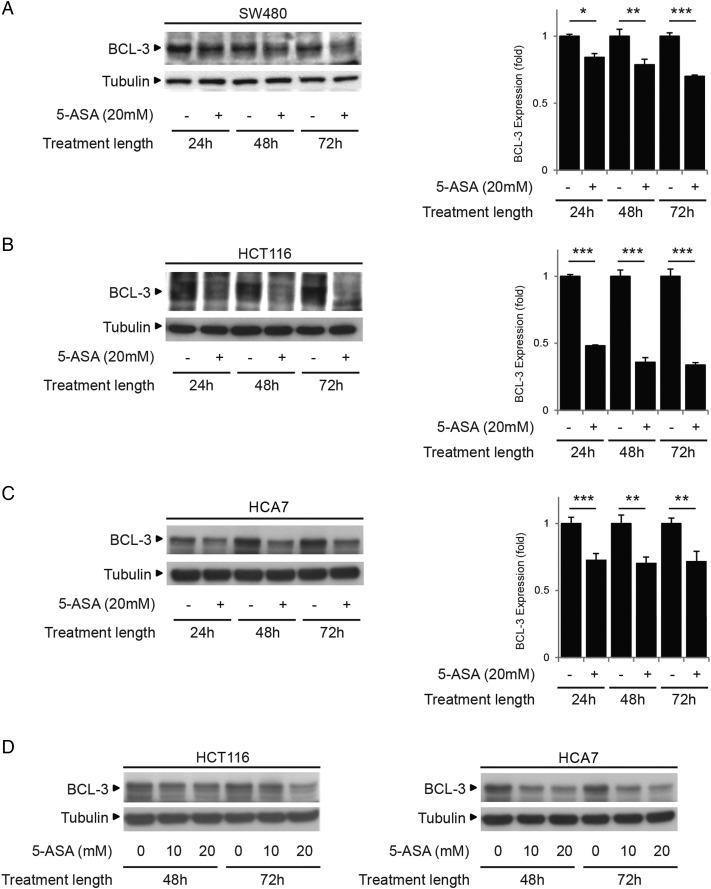
Results: We report increased BCL-3 expression in human colorectal cancers and demonstrate that BCL-3 expression promotes tumour cell survival in vitro and tumour growth in mouse xenografts in vivo, dependent on interaction with NF-κB p50 or p52 homodimers. We show that BCL-3 promotes cell survival under conditions relevant to the tumour microenvironment, protecting both colorectal adenoma and carcinoma cells from apoptosis via activation of the AKT survival pathway: AKT activation is mediated via both PI3K and mammalian target of rapamycin (mTOR) pathways, leading to phosphorylation of downstream targets GSK-3β and FoxO1/3a. Treatment with 5-ASA suppressed BCL-3 expression in colorectal cancer cells.
Conclusions: Our study helps to unravel the mechanism by which BCL-3 is linked to poor prognosis in colorectal cancer; we suggest that targeting BCL-3 activity represents an exciting therapeutic opportunity potentially increasing the sensitivity of tumour cells to conventional therapy.
Keywords: APOPTOSIS; CANCER PREVENTION; COLORECTAL CANCER; INFLAMMATION; NUCLEAR FACTOR KAPPA B.
Published by the BMJ Publishing Group Limited. For permission to use (where not already granted under a licence) please go to http://www.bmj.com/company/products-services/rights-and-licensing/
Figures
References
-
- Ferlay J, Soerjomataram I, Ervik M, et al. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 11, Colorectal Cancer [Internet]. Lyon, France: International Agency for Research on Cancer, 2013. http://globocan.iarc.fr/Pages/fact_sheets_cancer.aspx (accessed 07/08/2014).
-
- Cancer Research UK. Bowel cancer incidence report [Internet]. Cancer Research UK, 2014. http://www.cancerresearchuk.org/cancer-info/cancerstats/types/bowel/inci... (accessed 07/08/2014).
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous