

Multiple Interactions between Glucose and Brassinosteroid Signal Transduction Pathways in Arabidopsis Are Uncovered by Whole-Genome Transcriptional Profiling
- PMID: 26034265
- PMCID: PMC4741329
- DOI: 10.1104/pp.15.00495
Multiple Interactions between Glucose and Brassinosteroid Signal Transduction Pathways in Arabidopsis Are Uncovered by Whole-Genome Transcriptional Profiling
Abstract
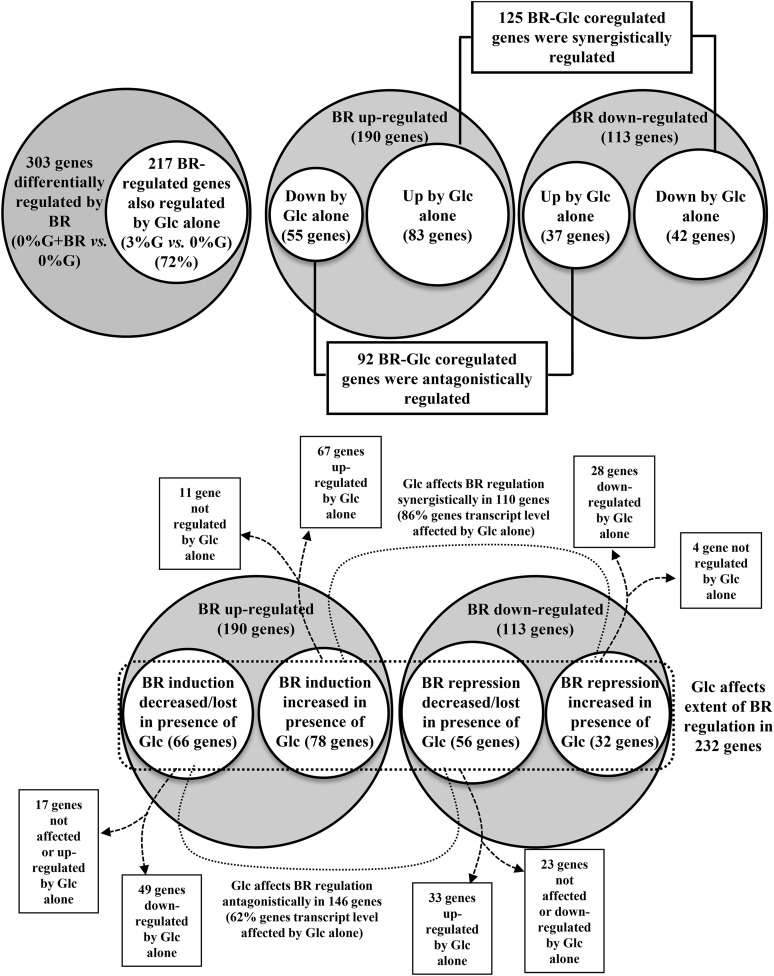
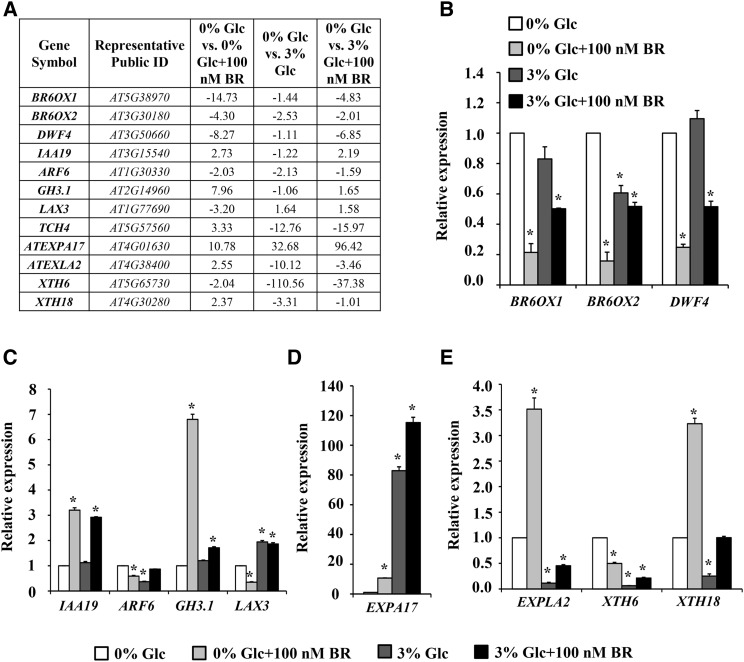
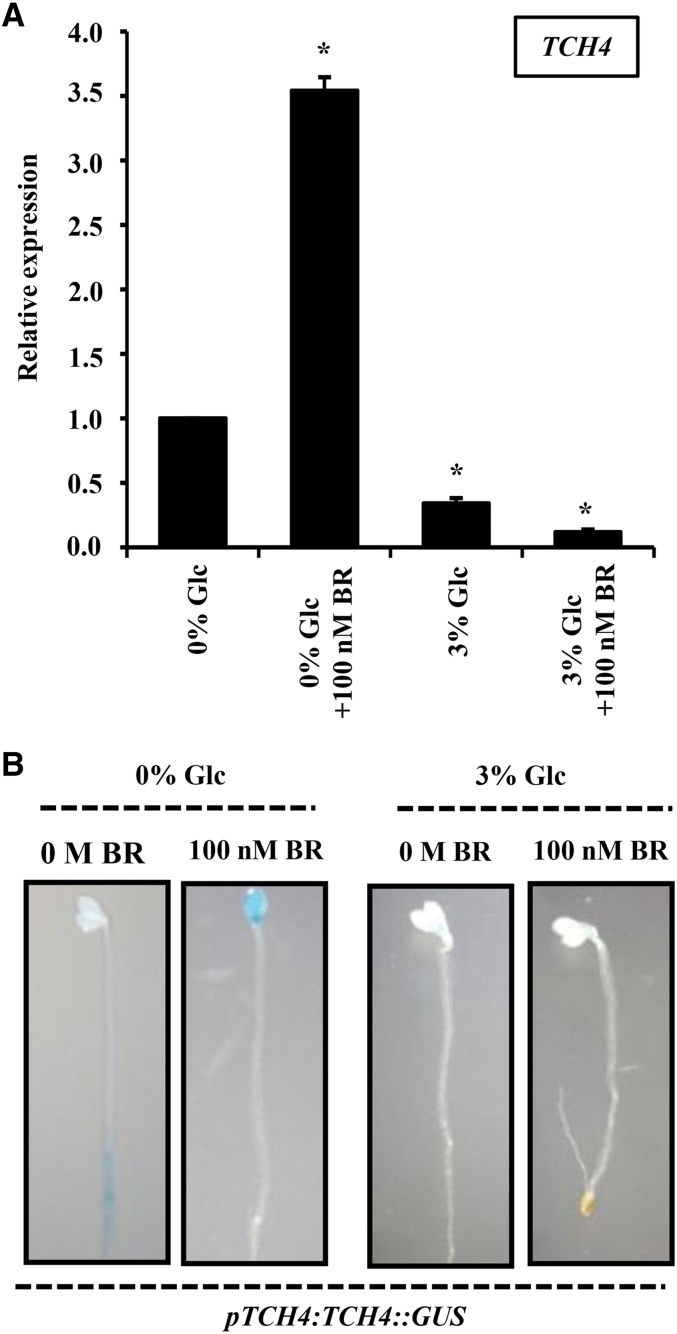
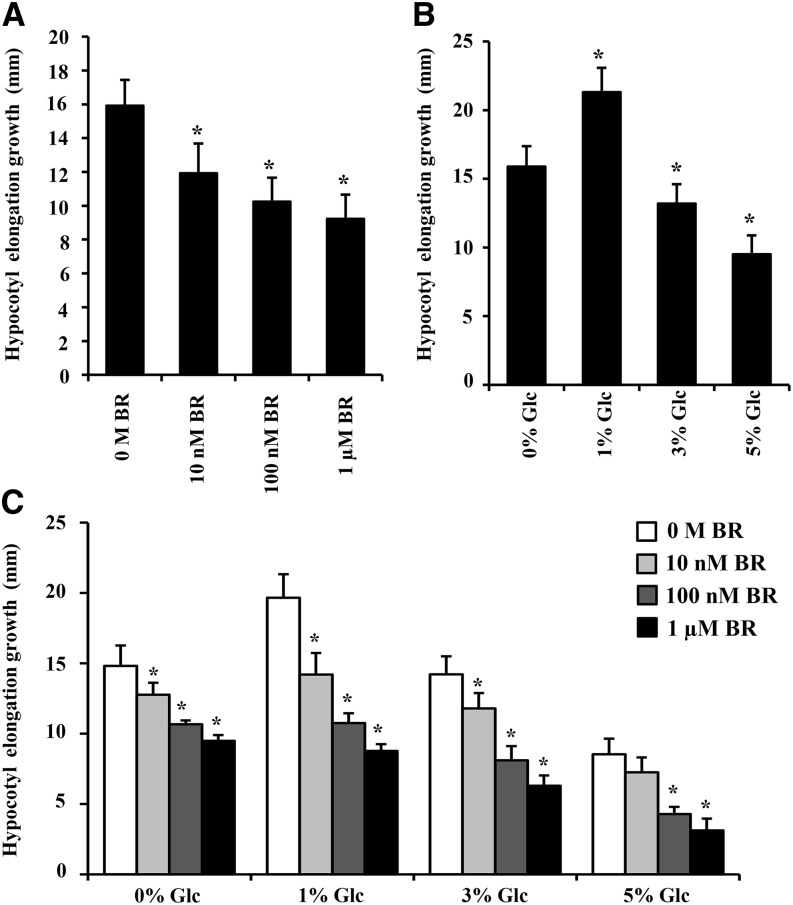
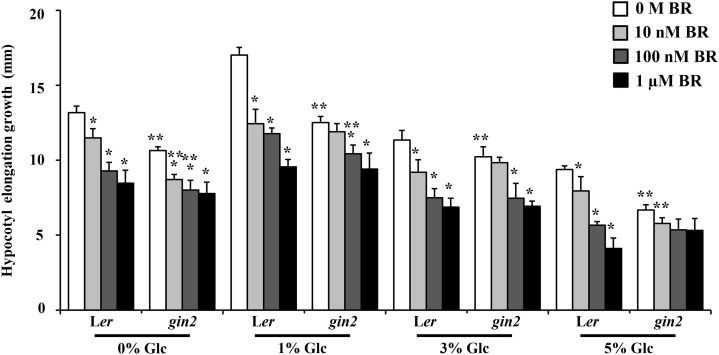
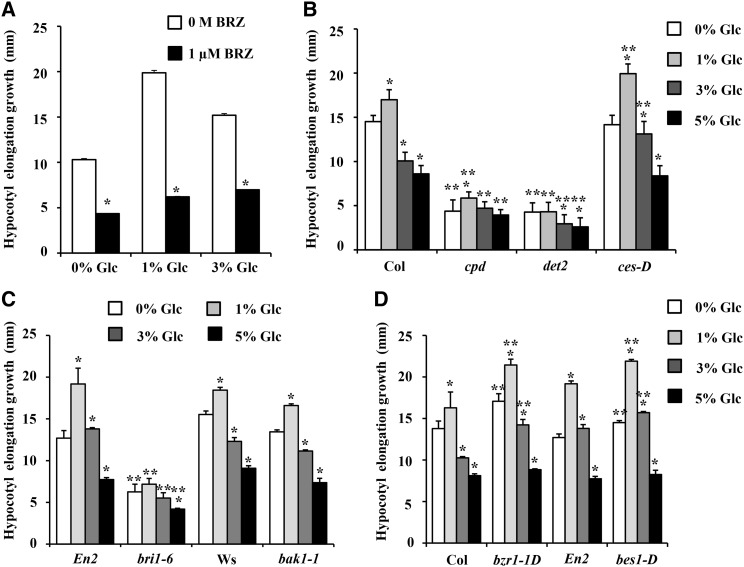
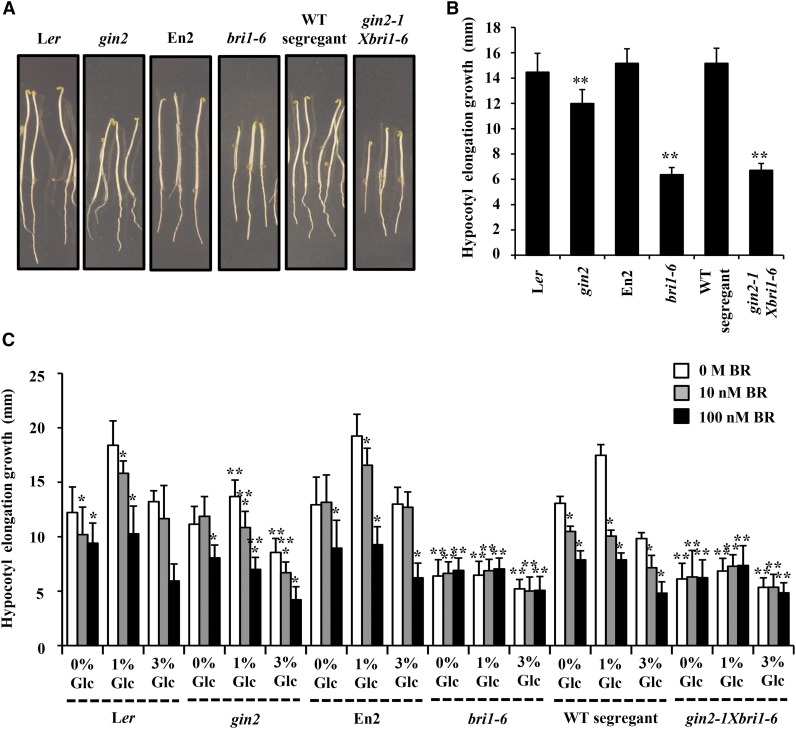
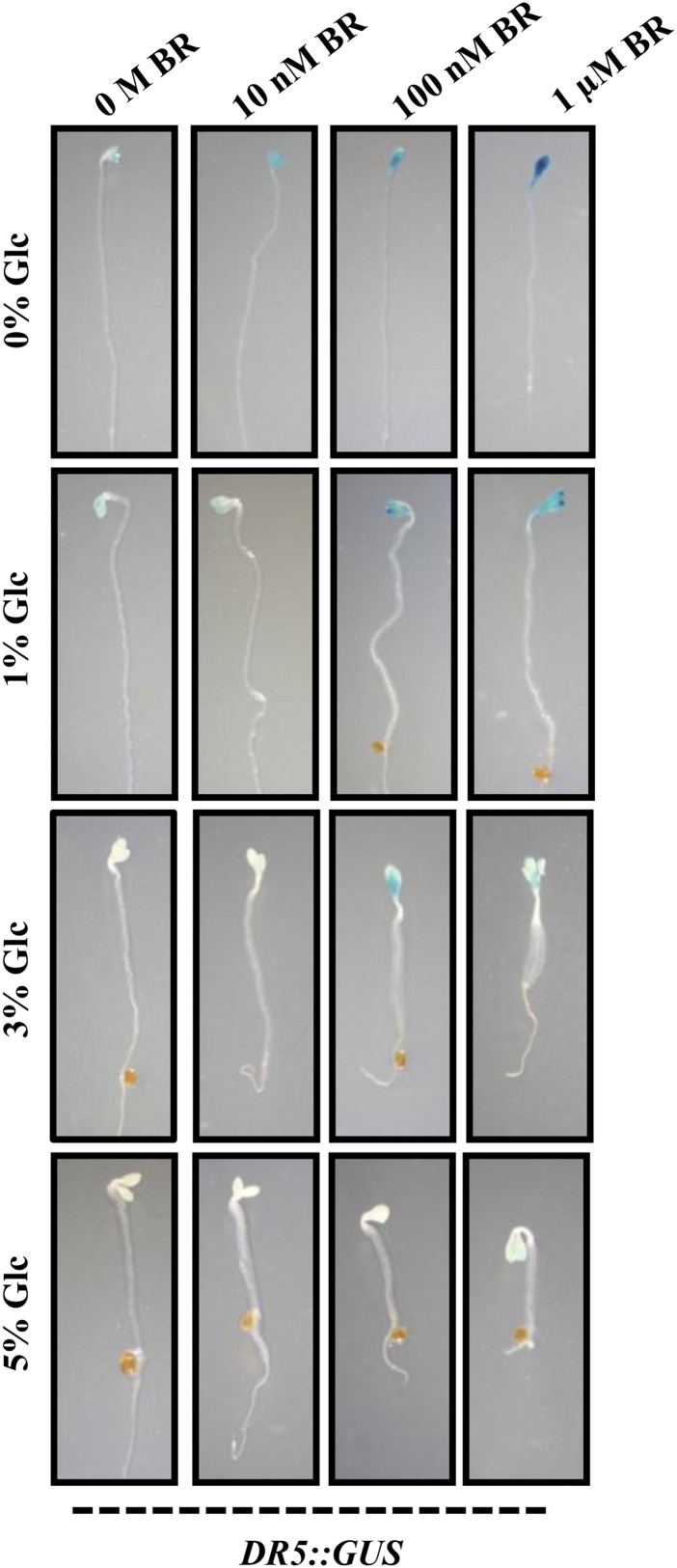
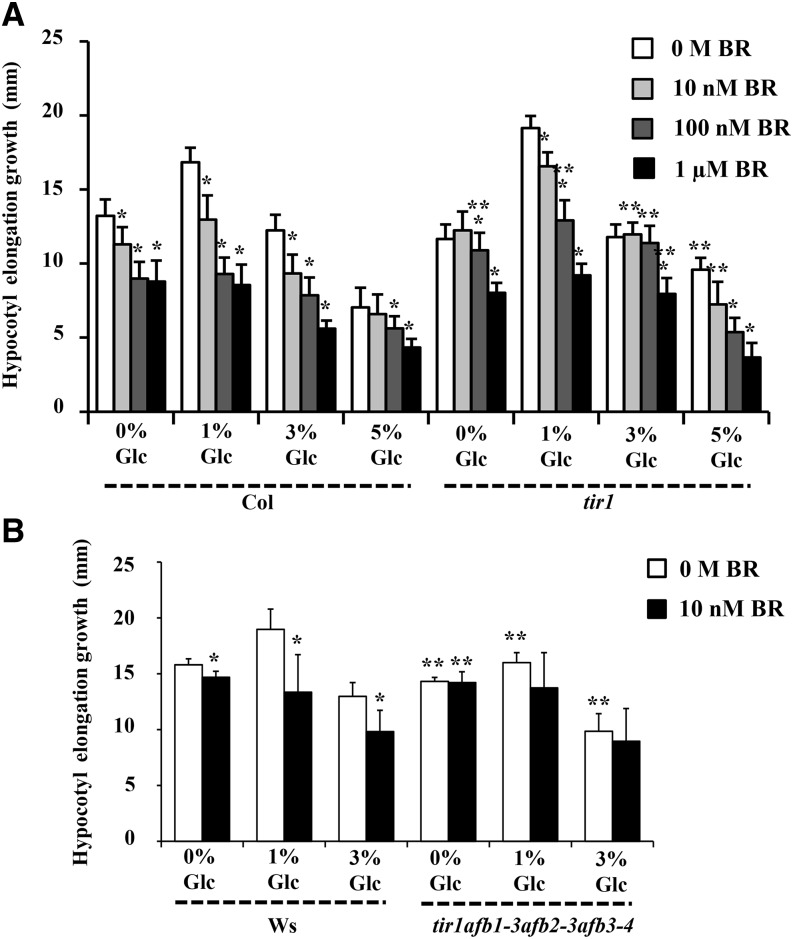
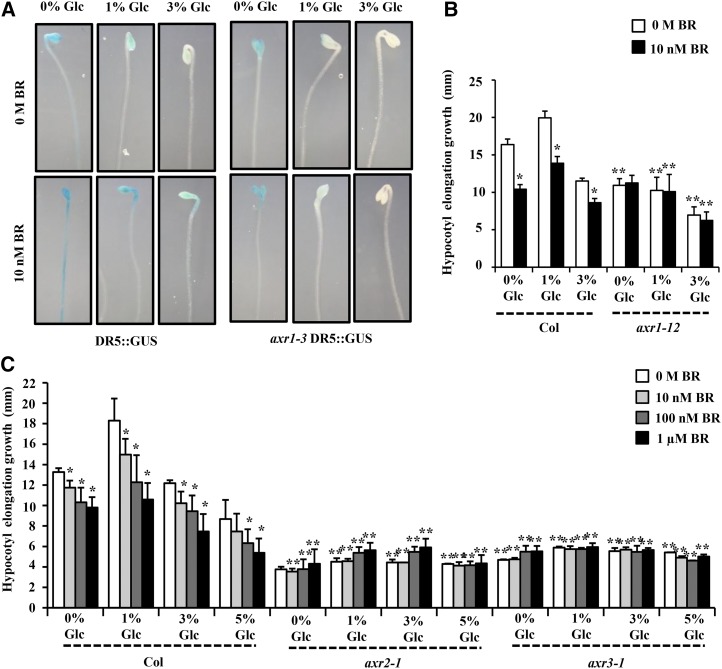
Brassinosteroid (BR) and glucose (Glc) regulate many common responses in plants. Here, we demonstrate that under etiolated growth conditions, extensive interdependence/overlap occurs between BR- and Glc-regulated gene expression as well as physiological responses. Glc could regulate the transcript level of 72% of BR-regulated genes at the whole-genome level, of which 58% of genes were affected synergistically and 42% of genes were regulated antagonistically. Presence of Glc along with BR in medium could affect BR induction/repression of 85% of BR-regulated genes. Glc could also regulate several genes involved in BR metabolism and signaling. Both BR and Glc coregulate a large number of genes involved in abiotic/biotic stress responses and growth and development. Physiologically, Glc and BR interact to regulate hypocotyl elongation growth of etiolated Arabidopsis (Arabidopsis thaliana) seedlings in a dose-dependent manner. Glc may interact with BR via a hexokinase1 (HXK1)-mediated pathway to regulate etiolated hypocotyl elongation. Brassinosteroid insensitive1 (BRI1) is epistatic to HXK1, as the Glc insensitive2bri1-6 double mutant displayed severe defects in hypocotyl elongation growth similar to its bri1-6 parent. Analysis of Glc and BR sensitivity in mutants defective in auxin response/signaling further suggested that Glc and BR signals may converge at S-phase kinase-associated protein1-Cullin-F-box-transport inhibitor response1/auxin-related f-box-auxin/indole-3-acetic acid-mediated auxin-signaling machinery to regulate etiolated hypocotyl elongation growth in Arabidopsis.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures
Similar articles
-
Interaction between glucose and brassinosteroid during the regulation of lateral root development in Arabidopsis.Plant Physiol. 2015 May;168(1):307-20. doi: 10.1104/pp.114.256313. Epub 2015 Mar 25. Plant Physiol. 2015. PMID: 25810094 Free PMC article.
-
Brassinosteroids regulate the differential growth of Arabidopsis hypocotyls through auxin signaling components IAA19 and ARF7.Mol Plant. 2013 May;6(3):887-904. doi: 10.1093/mp/sss123. Epub 2012 Nov 2. Mol Plant. 2013. PMID: 23125315
-
Brassinosteroid signaling converges with SUPPRESSOR OF PHYTOCHROME B4-#3 to influence the expression of SMALL AUXIN UP RNA genes and hypocotyl growth.Plant J. 2017 Mar;89(6):1133-1145. doi: 10.1111/tpj.13451. Epub 2017 Feb 17. Plant J. 2017. PMID: 27984677 Free PMC article.
-
The physiological and molecular mechanism of brassinosteroid in response to stress: a review.Biol Res. 2018 Nov 12;51(1):46. doi: 10.1186/s40659-018-0195-2. Biol Res. 2018. PMID: 30419959 Free PMC article. Review.
-
Brassinosteroid signaling in plant development and adaptation to stress.Development. 2019 Mar 14;146(5):dev151894. doi: 10.1242/dev.151894. Development. 2019. PMID: 30872266 Free PMC article. Review.
Cited by
-
Molecular mechanisms of resistance to Myzus persicae conferred by the peach Rm2 gene: A multi-omics view.Front Plant Sci. 2022 Oct 5;13:992544. doi: 10.3389/fpls.2022.992544. eCollection 2022. Front Plant Sci. 2022. PMID: 36275570 Free PMC article.
-
Phosphatidic Acid in Plant Hormonal Signaling: From Target Proteins to Membrane Conformations.Int J Mol Sci. 2022 Mar 17;23(6):3227. doi: 10.3390/ijms23063227. Int J Mol Sci. 2022. PMID: 35328648 Free PMC article. Review.
-
Arabidopsis Target of Rapamycin Coordinates With Transcriptional and Epigenetic Machinery to Regulate Thermotolerance.Front Plant Sci. 2021 Oct 28;12:741965. doi: 10.3389/fpls.2021.741965. eCollection 2021. Front Plant Sci. 2021. PMID: 34777423 Free PMC article.
-
Increasing Sugar Content in Source for Biofuel Production Using Agrochemical and Genetic Approaches at the Stages of BioMass Preharvesting and Harvesting.Molecules. 2022 Aug 16;27(16):5210. doi: 10.3390/molecules27165210. Molecules. 2022. PMID: 36014450 Free PMC article. Review.
-
Arabidopsis RSS1 Mediates Cross-Talk Between Glucose and Light Signaling During Hypocotyl Elongation Growth.Sci Rep. 2017 Nov 23;7(1):16101. doi: 10.1038/s41598-017-16239-y. Sci Rep. 2017. PMID: 29170398 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
