

Convergence of eicosanoid and integrin biology: 12-lipoxygenase seeks a partner
- PMID: 26037302
- PMCID: PMC4453211
- DOI: 10.1186/s12943-015-0382-5
Convergence of eicosanoid and integrin biology: 12-lipoxygenase seeks a partner
Abstract
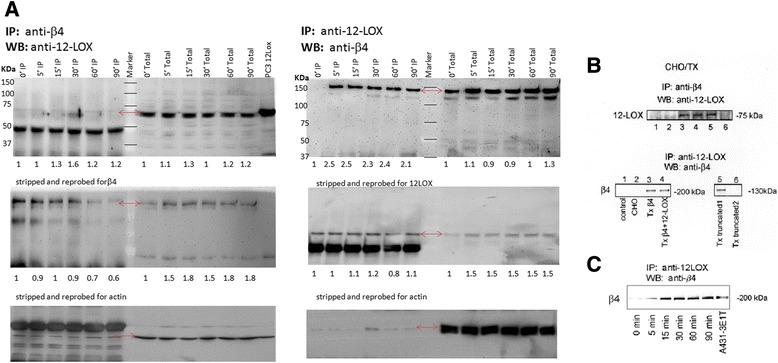
Background: Integrins and enzymes of the eicosanoid pathway are both well-established contributors to cancer. However, this is the first report of the interdependence of the two signaling systems. In a screen for proteins that interacted with, and thereby potentially regulated, the human platelet-type 12-lipoxygenase (12-LOX, ALOX12), we identified the integrin β4 (ITGB4).
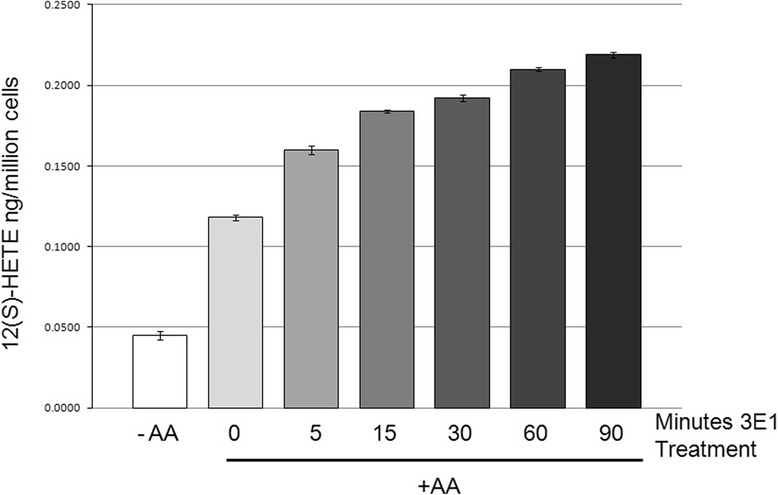
Methods: Using a cultured mammalian cell model, we have demonstrated that ITGB4 stimulation leads to recruitment of 12-LOX from the cytosol to the membrane where it physically interacts with the integrin to become enzymatically active to produce 12(S)-HETE, a known bioactive lipid metabolite that regulates numerous cancer phenotypes.
Results: The net effect of the interaction was the prevention of cell death in response to starvation. Additionally, regulation of β4-mediated, EGF-stimulated invasion was shown to be dependent on 12-LOX, and downstream Erk signaling in response to ITGB4 activation also required 12-LOX.
Conclusions: This is the first report of an enzyme of the eicosanoid pathway being recruited to and regulated by activated β4 integrin. Integrin β4 has recently been shown to induce expansion of prostate tumor progenitors and there is a strong correlation between stage/grade of prostate cancer and 12-LOX expression. The 12-LOX enzymatic product, 12(S)-HETE, regulates angiogenesis and cell migration in many cancer types. Therefore, disruption of integrin β4-12LOX interaction could reduce the pro-inflammatory oncogenic activity of 12-LOX. This report on the consequences of 12-LOX and ITGB4 interaction sets a precedent for the linkage of integrin and eicosanoid biology through direct protein-protein association.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources