

The Rift Valley fever accessory proteins NSm and P78/NSm-GN are distinct determinants of virus propagation in vertebrate and invertebrate hosts
- PMID: 26038497
- PMCID: PMC4217093
- DOI: 10.1038/emi.2014.71
The Rift Valley fever accessory proteins NSm and P78/NSm-GN are distinct determinants of virus propagation in vertebrate and invertebrate hosts
Abstract
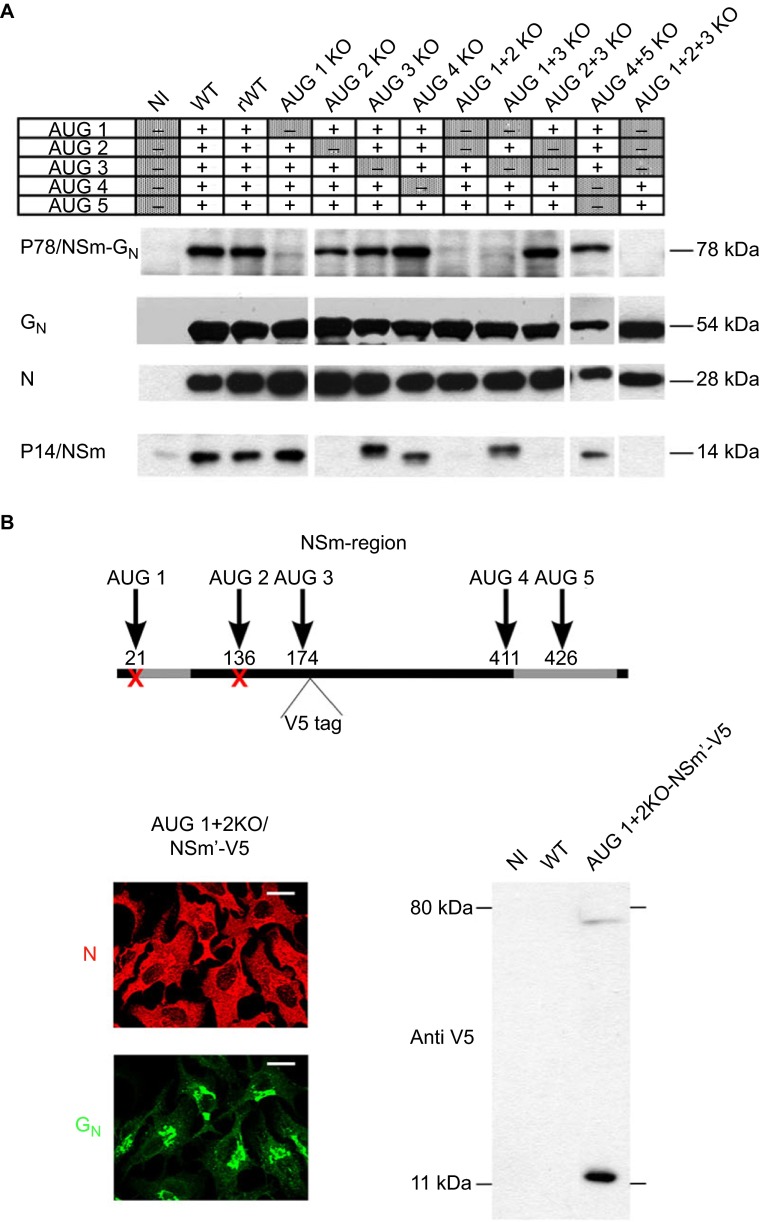
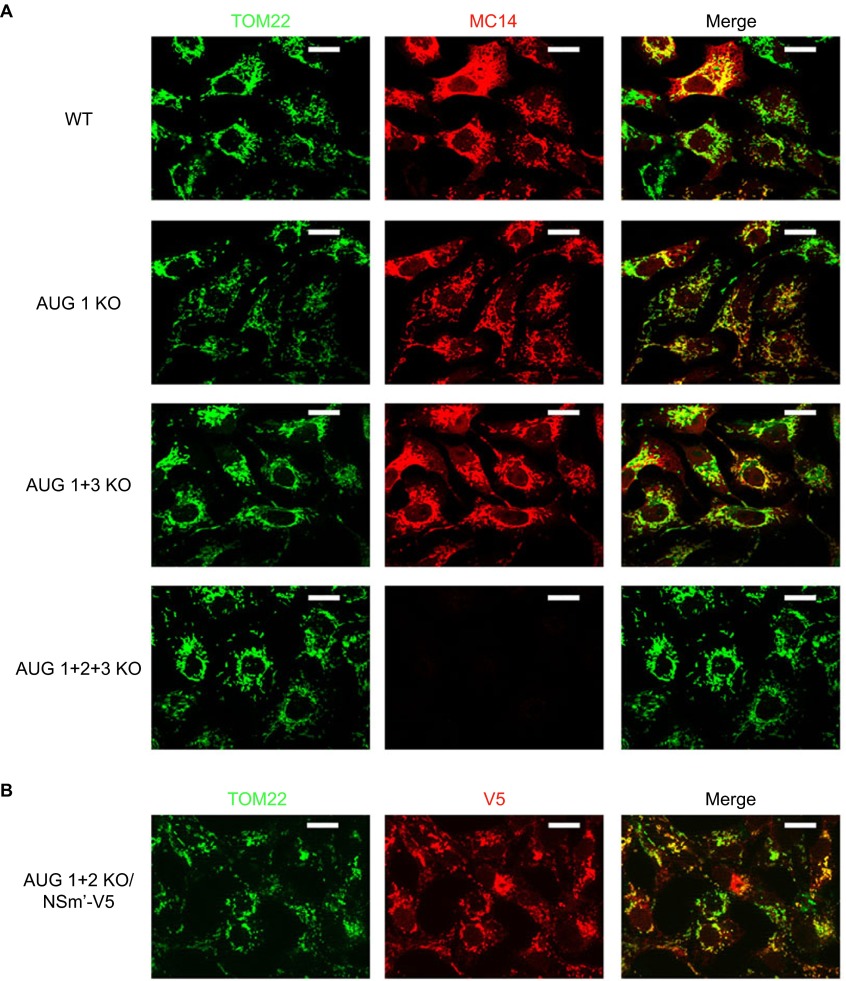
Rift Valley fever virus (RVFV) is an enzootic virus circulating in Africa that is transmitted to its vertebrate host by a mosquito vector and causes severe clinical manifestations in humans and ruminants. RVFV has a tripartite genome of negative or ambisense polarity. The M segment contains five in-frame AUG codons that are alternatively used for the synthesis of two major structural glycoproteins, GN and GC, and at least two accessory proteins, NSm, a 14-kDa cytosolic protein, and P78/NSm-GN, a 78-kDa glycoprotein. To determine the relative contribution of P78 and NSm to RVFV infectivity, AUG codons were knocked out to generate mutant viruses expressing various sets of the M-encoded proteins. We found that, in the absence of the second AUG codon used to express NSm, a 13-kDa protein corresponding to an N-terminally truncated form of NSm, named NSm', was synthesized from AUG 3. None of the individual accessory proteins had any significant impact on RVFV virulence in mice. However, a mutant virus lacking both NSm and NSm' was strongly attenuated in mice and grew to reduced titers in murine macrophages, a major target cell type of RVFV. In contrast, P78 was not associated with reduced viral virulence in mice, yet it appeared as a major determinant of virus dissemination in mosquitoes. This study demonstrates how related accessory proteins differentially contribute to RVFV propagation in mammalian and arthropod hosts.
Keywords: arbovirus; attenuation; bunyavirus; minor structural protein; non-structural protein; transmission; vector; virulence factor.
Figures



References
-
- Daubney R, Hudson JR, Garnham PC. Enzootic hepatitis or rift valley fever. An undescribed virus disease of sheep cattle and man from east africa. J Pathol Bacteriol. 1931;34:545–579.
-
- Findlay GM. Rift valley fever or enzootic hepatitis. Trans R Soc Trop Med Hyg. 1932;25:229–IN211.
-
- Bird BH, Ksiazek TG, Nichol ST, Maclachlan NJ. Rift Valley fever virus. J Am Vet Med Assoc. 2009;234:883–893. - PubMed
-
- Meegan JM, Hoogstraal H, Moussa MI. An epizootic of Rift Valley fever in Egypt in 1977. Vet Rec. 1979;105:124–125. - PubMed
-
- Arthur RR, El-Sharkawy MS, Cope SE, et al. Recurrence of Rift Valley fever in Egypt. Lancet. 1993;342:1149–1150. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous