

Pathobiological features of a novel, highly pathogenic avian influenza A(H5N8) virus
- PMID: 26038499
- PMCID: PMC4217095
- DOI: 10.1038/emi.2014.75
Pathobiological features of a novel, highly pathogenic avian influenza A(H5N8) virus
Abstract
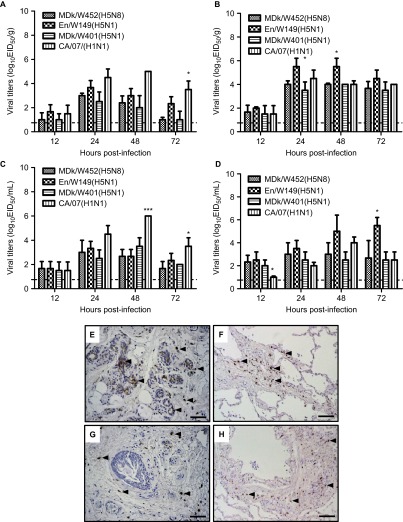
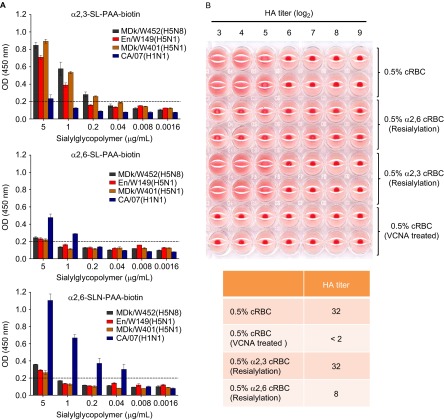
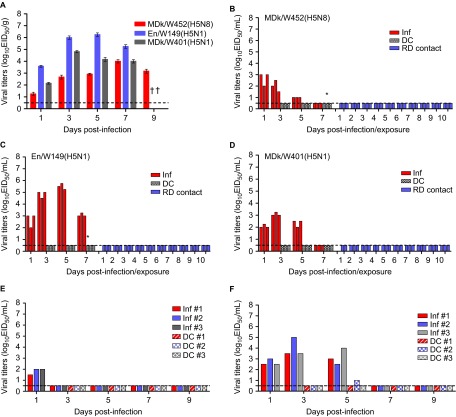
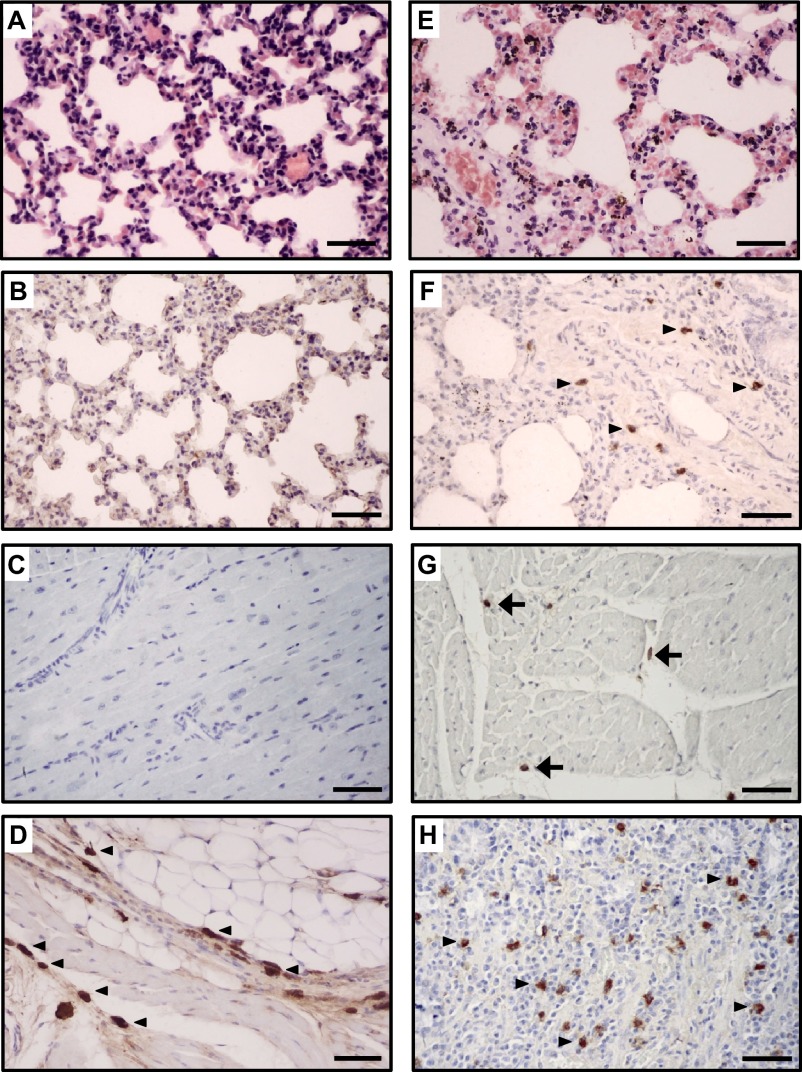
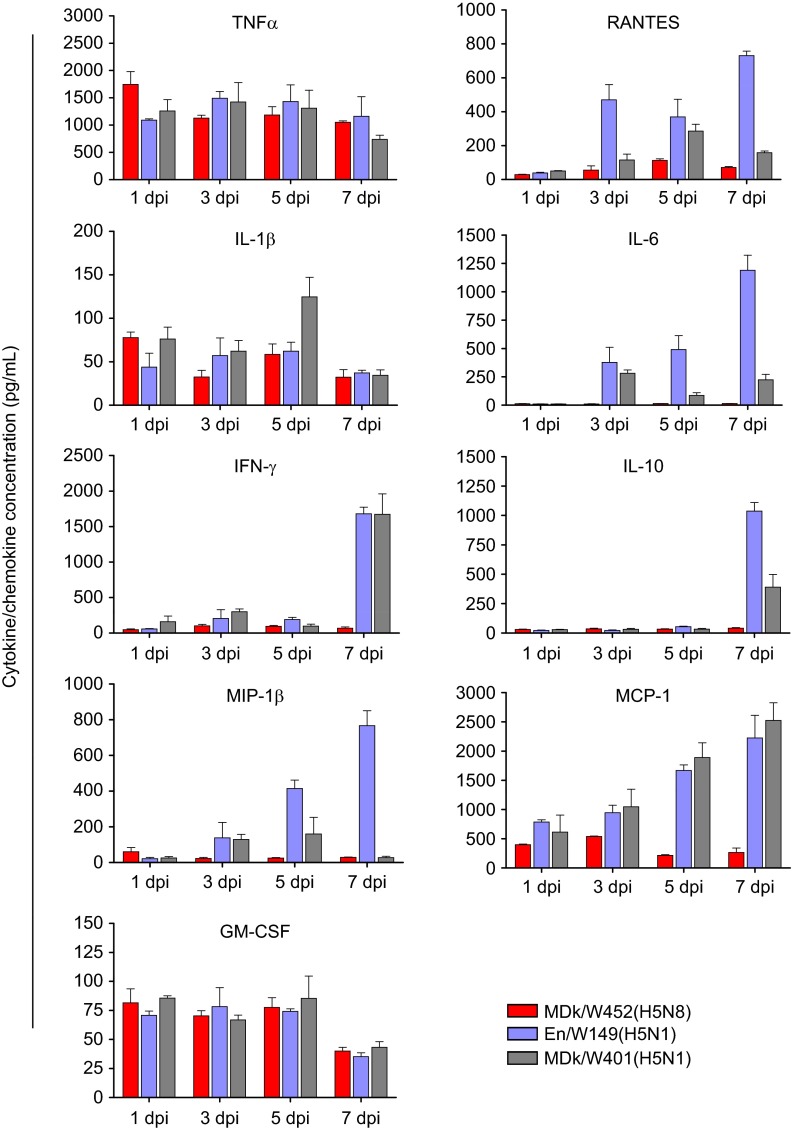
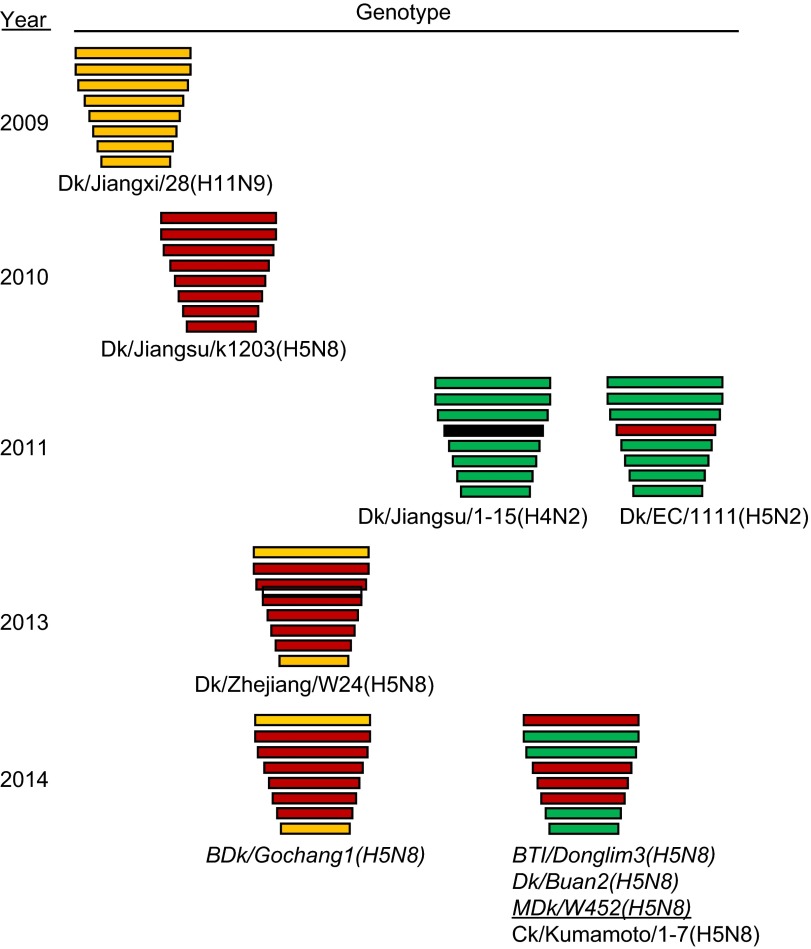
The endemicity of highly pathogenic avian influenza (HPAI) A(H5N1) viruses in Asia has led to the generation of reassortant H5 strains with novel gene constellations. A newly emerged HPAI A(H5N8) virus caused poultry outbreaks in the Republic of Korea in 2014. Because newly emerging high-pathogenicity H5 viruses continue to pose public health risks, it is imperative that their pathobiological properties be examined. Here, we characterized A/mallard duck/Korea/W452/2014 (MDk/W452(H5N8)), a representative virus, and evaluated its pathogenic and pandemic potential in various animal models. We found that MDk/W452(H5N8), which originated from the reassortment of wild bird viruses harbored by migratory waterfowl in eastern China, replicated systemically and was lethal in chickens, but appeared to be attenuated, albeit efficiently transmitted, in ducks. Despite predominant attachment to avian-like virus receptors, MDk/W452(H5N8) also exhibited detectable human virus-like receptor binding and replicated in human respiratory tract tissues. In mice, MDk/W452(H5N8) was moderately pathogenic and had limited tissue tropism relative to previous HPAI A(H5N1) viruses. It also induced moderate nasal wash titers in inoculated ferrets; additionally, it was recovered in extrapulmonary tissues and one of three direct-contact ferrets seroconverted without shedding. Moreover, domesticated cats appeared to be more susceptible than dogs to virus infection. With their potential to become established in ducks, continued circulation of A(H5N8) viruses could alter the genetic evolution of pre-existing avian poultry strains. Overall, detailed virological investigation remains a necessity given the capacity of H5 viruses to evolve to cause human illness with few changes in the viral genome.
Keywords: HPAI A(H5N8); avian influenza virus; genetic evolution; migratory waterfowl; reassortment.
Figures
References
-
- Eagles D, Siregar ES, Dung DH, Weaver J, Wong F, Daniels P. H5N1 highly pathogenic avian influenza in Southeast Asia. Rev Sci Tech. 2009;28:341–348. - PubMed
-
- World Health Organization/Global Influenza Programme Cumulative number of confirmed human cases for avian influenza A(H5N1) reported to WHO, 2003–2014 Geneva: WHO; 2014. Available at http://www.who.int/influenza/human_animal_interface/EN_GIP_20140124Cumul... (accessed 24 January 2014).
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous