

Arterivirus nsp12 versus the coronavirus nsp16 2'-O-methyltransferase: comparison of the C-terminal cleavage products of two nidovirus pp1ab polyproteins
- PMID: 26041874
- PMCID: PMC7081073
- DOI: 10.1099/vir.0.000209
Arterivirus nsp12 versus the coronavirus nsp16 2'-O-methyltransferase: comparison of the C-terminal cleavage products of two nidovirus pp1ab polyproteins
Abstract
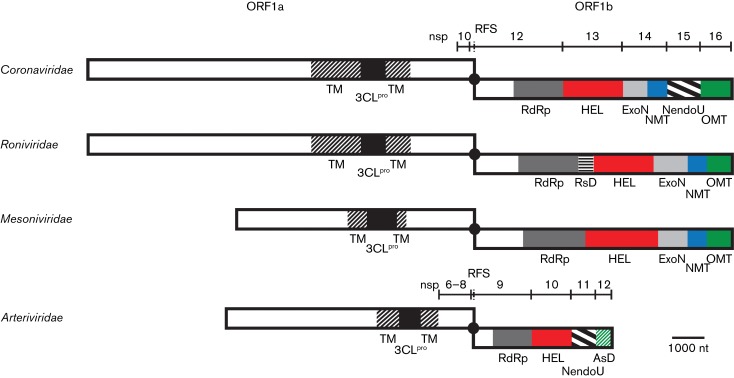
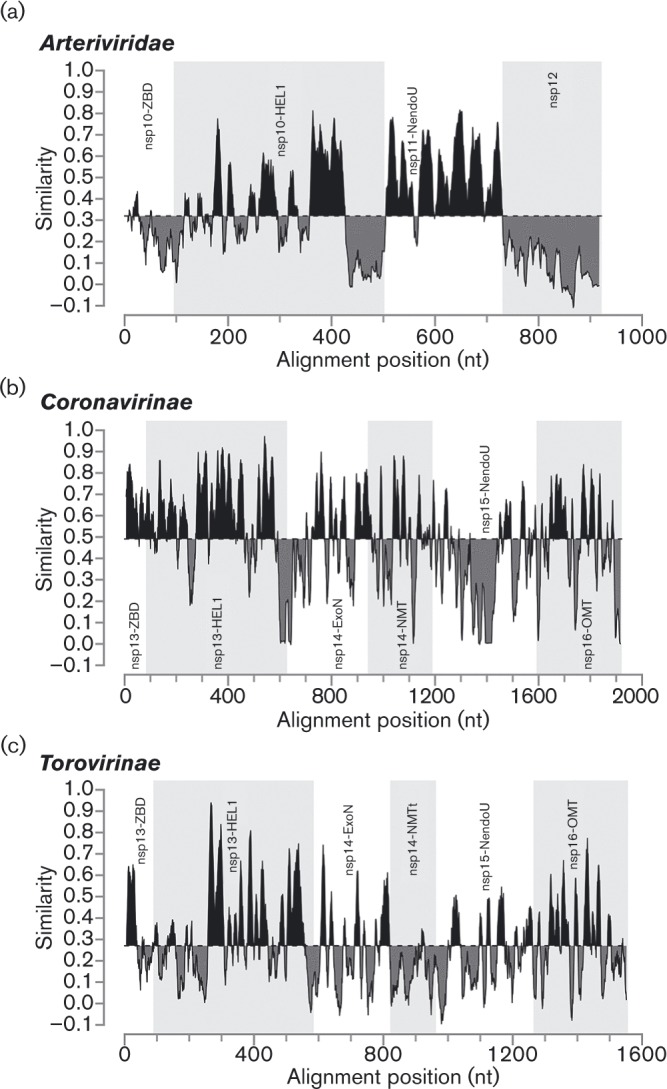
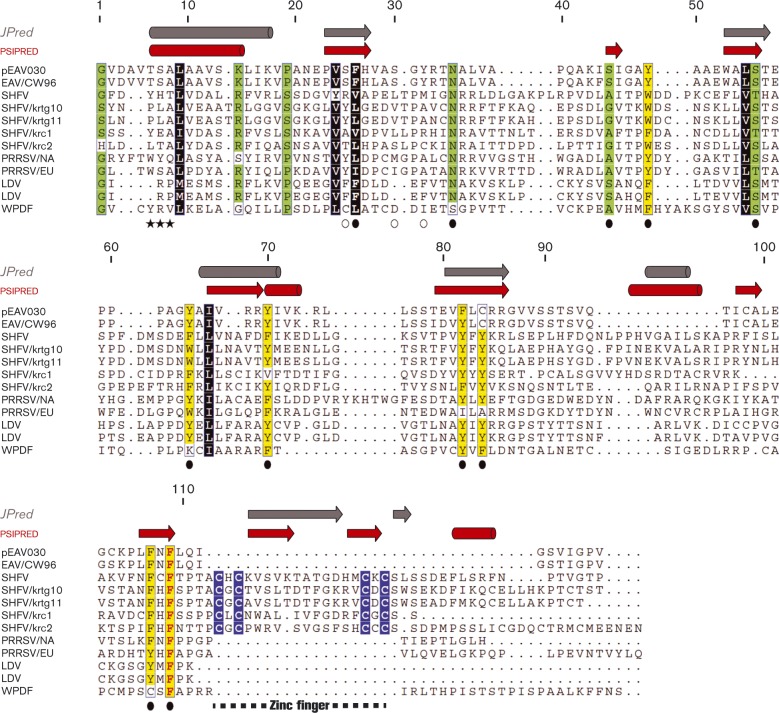
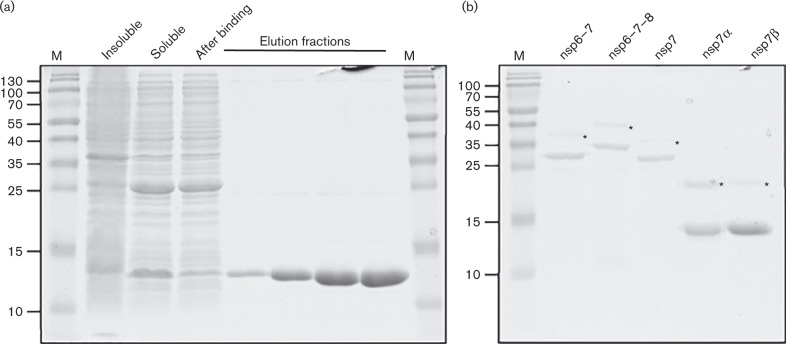
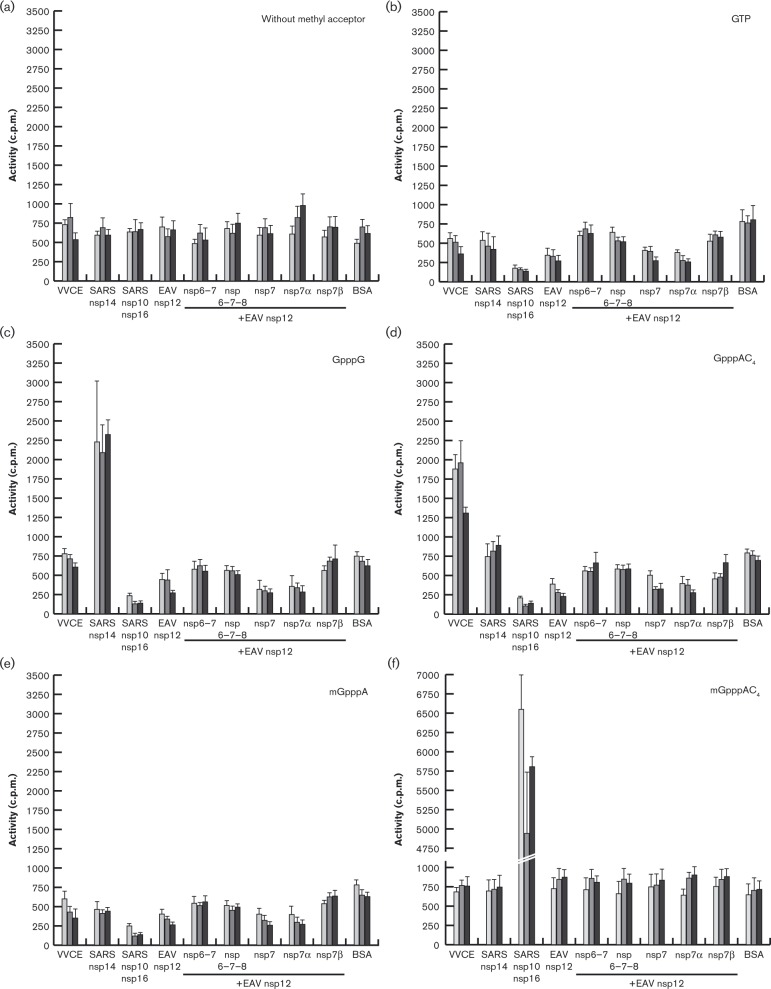
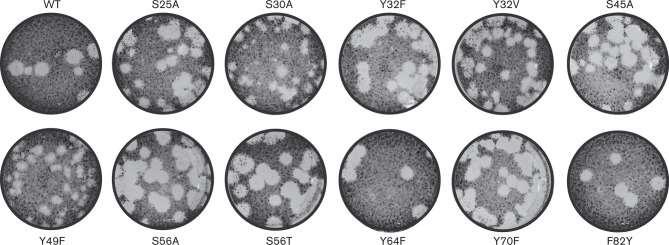
The 3'-terminal domain of the most conserved ORF1b in three of the four families of the order Nidovirales (except for the family Arteriviridae) encodes a (putative) 2'-O-methyltransferase (2'-O-MTase), known as non structural protein (nsp) 16 in the family Coronaviridae and implicated in methylation of the 5' cap structure of nidoviral mRNAs. As with coronavirus transcripts, arterivirus mRNAs are assumed to possess a 5' cap although no candidate MTases have been identified thus far. To address this knowledge gap, we analysed the uncharacterized nsp12 of arteriviruses, which occupies the ORF1b position equivalent to that of the nidovirus 2'-O-MTase (coronavirus nsp16). In our in-depth bioinformatics analysis of nsp12, the protein was confirmed to be family specific whilst having diverged much further than other nidovirus ORF1b-encoded proteins, including those of the family Coronaviridae. Only one invariant and several partially conserved, predominantly aromatic residues were identified in nsp12, which may adopt a structure with alternating α-helices and β-strands, an organization also found in known MTases. However, no statistically significant similarity was found between nsp12 and the twofold larger coronavirus nsp16, nor could we detect MTase activity in biochemical assays using recombinant equine arteritis virus (EAV) nsp12. Our further analysis established that this subunit is essential for replication of this prototypic arterivirus. Using reverse genetics, we assessed the impact of 25 substitutions at 14 positions, yielding virus phenotypes ranging from WT-like to non-viable. Notably, replacement of the invariant phenylalanine 109 with tyrosine was lethal. We concluded that nsp12 plays an essential role during EAV replication, possibly by acting as a co-factor for another enzyme.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous