

Cyclic Regulation of Sensory Perception by a Female Hormone Alters Behavior
- PMID: 26046438
- PMCID: PMC4501503
- DOI: 10.1016/j.cell.2015.04.052
Cyclic Regulation of Sensory Perception by a Female Hormone Alters Behavior
Abstract
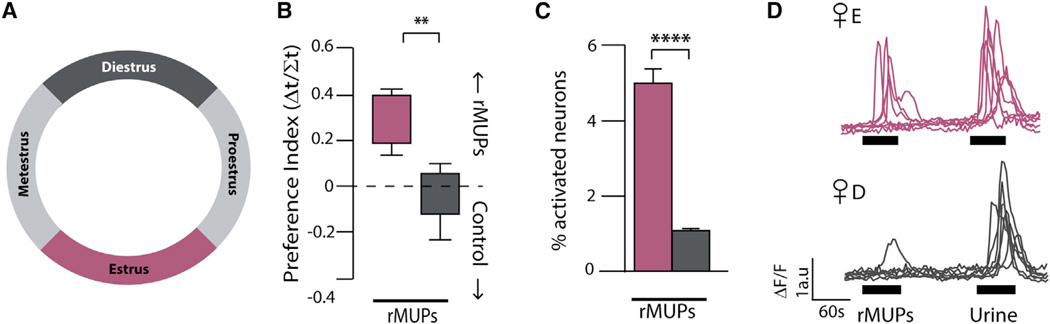
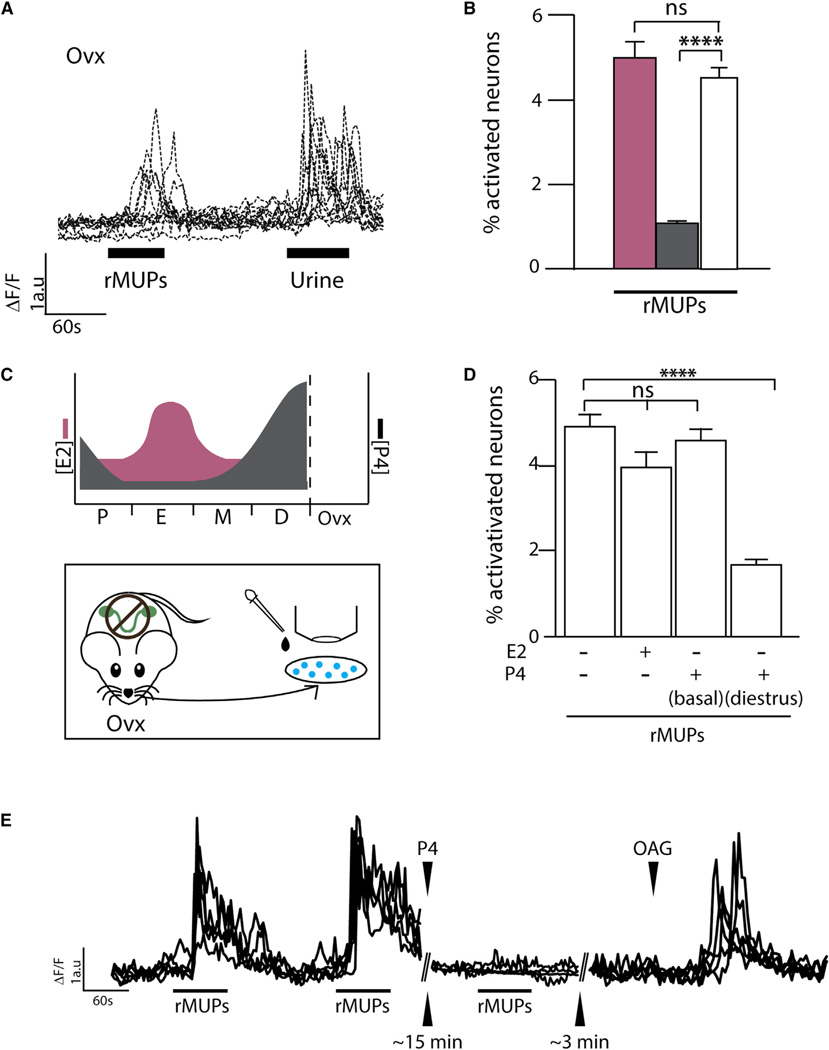
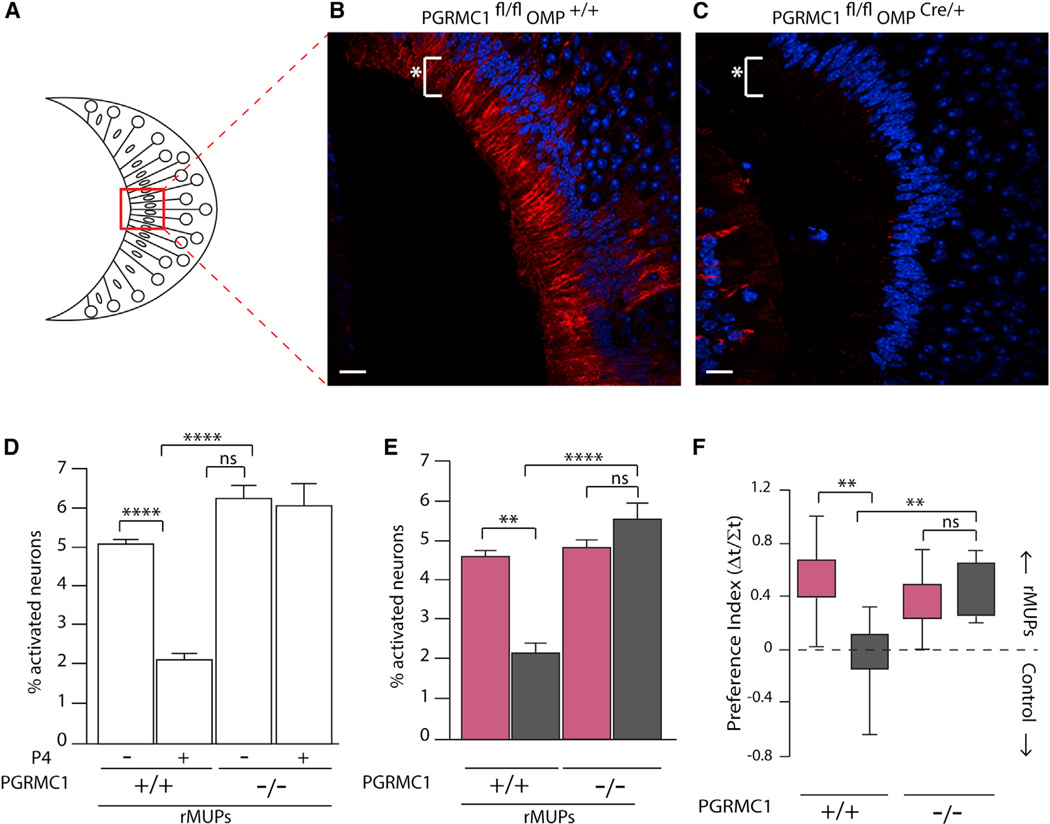
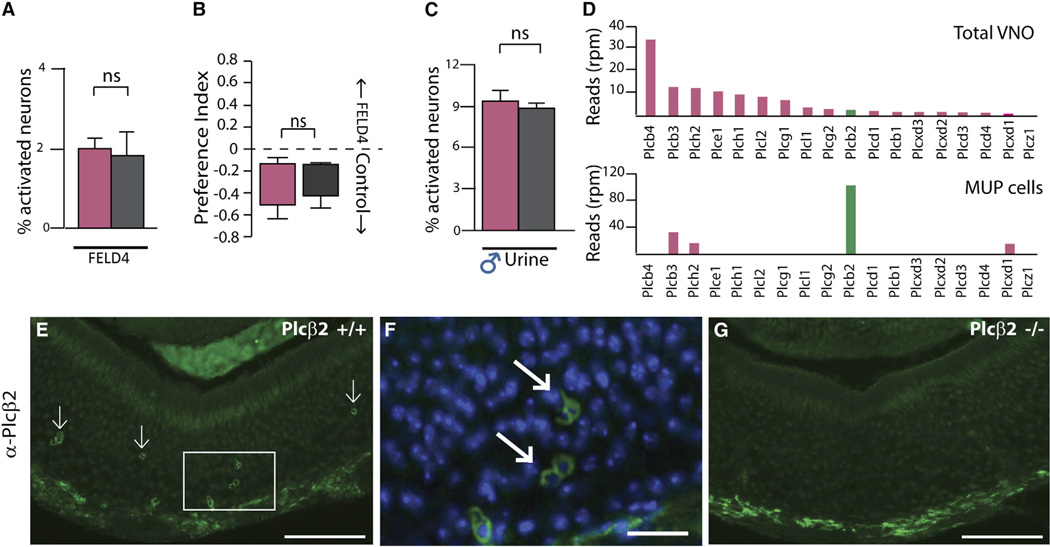
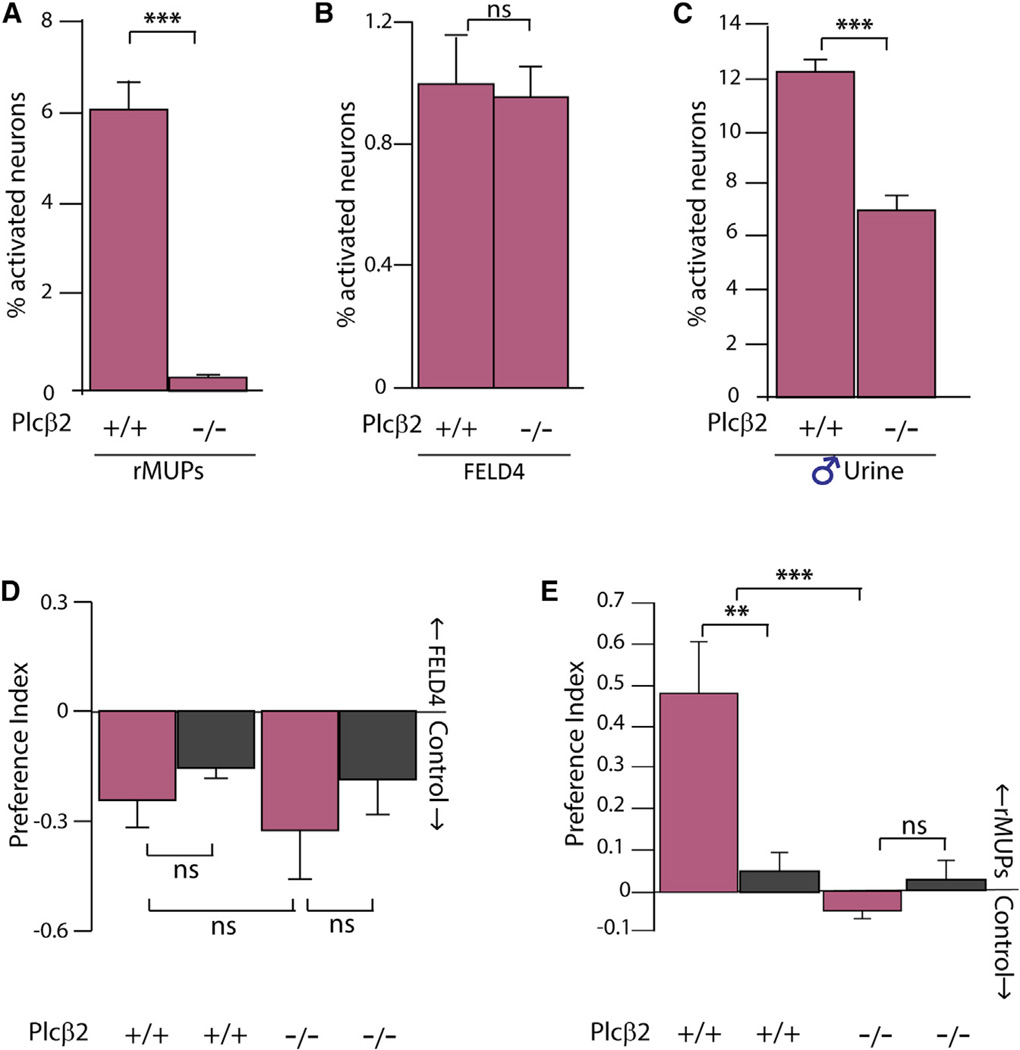
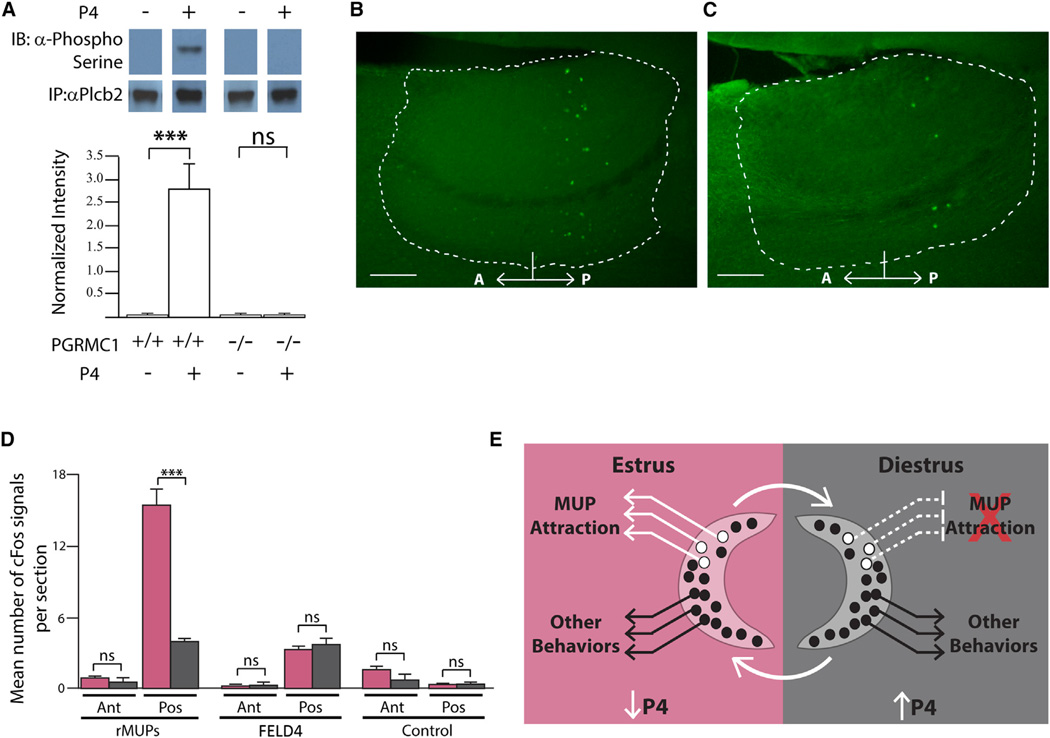
Females may display dramatically different behavior depending on their state of ovulation. This is thought to occur through sex-specific hormones acting on behavioral centers in the brain. Whether incoming sensory activity also differs across the ovulation cycle to alter behavior has not been investigated. Here, we show that female mouse vomeronasal sensory neurons (VSNs) are temporarily and specifically rendered "blind" to a subset of male-emitted pheromone ligands during diestrus yet fully detect and respond to the same ligands during estrus. VSN silencing occurs through the action of the female sex-steroid progesterone. Not all VSNs are targeted for silencing; those detecting cat ligands remain continuously active irrespective of the estrous state. We identify the signaling components that account for the capacity of progesterone to target specific subsets of male-pheromone responsive neurons for inactivation. These findings indicate that internal physiology can selectively and directly modulate sensory input to produce state-specific behavior. PAPERCLIP.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Chamero P, Marton TF, Logan DW, Flanagan K, Cruz JR, Saghatelian A, Cravatt BF, Stowers L. Identification of protein pheromones that promote aggressive behaviour. Nature. 2007;450:899–902. - PubMed
-
- Chamero P, Leinders-Zufall T, Zufall F. From genes to social communication: molecular sensing by the vomeronasal organ. Trends Neurosci. 2012;35:597–606. - PubMed
-
- Dulac O, Torello AT. Molecular detection of pheromone signals in mammals: from genes to behaviour. Nat. Rev. Neurosci. 2003;4:551–562. - PubMed
-
- Eggan K, Baldwin K, Tackett M, Osborne J, Gogos J, Chess A, Axel R, Jaenisch R. Mice cloned from olfactory sensory neurons. Nature. 2004;428:44–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- DC009413/DC/NIDCD NIH HHS/United States
- T32 HD040372/HD/NICHD NIH HHS/United States
- T32 GM007754/GM/NIGMS NIH HHS/United States
- R01 DC009413/DC/NIDCD NIH HHS/United States
- R01 DC012095/DC/NIDCD NIH HHS/United States
- R01 DC006885/DC/NIDCD NIH HHS/United States
- DC010857/DC/NIDCD NIH HHS/United States
- R01 DC010857/DC/NIDCD NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R21 RR030264/RR/NCRR NIH HHS/United States
- DC012095/DC/NIDCD NIH HHS/United States
- DC006885/DC/NIDCD NIH HHS/United States
- 098051/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
