

Transgenic songbirds with suppressed or enhanced activity of CREB transcription factor
- PMID: 26048905
- PMCID: PMC4475973
- DOI: 10.1073/pnas.1413484112
Transgenic songbirds with suppressed or enhanced activity of CREB transcription factor
Abstract
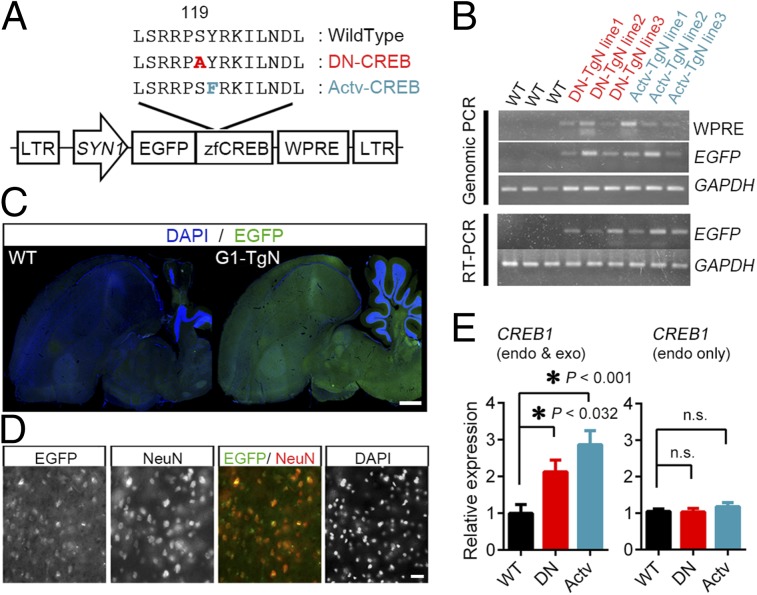
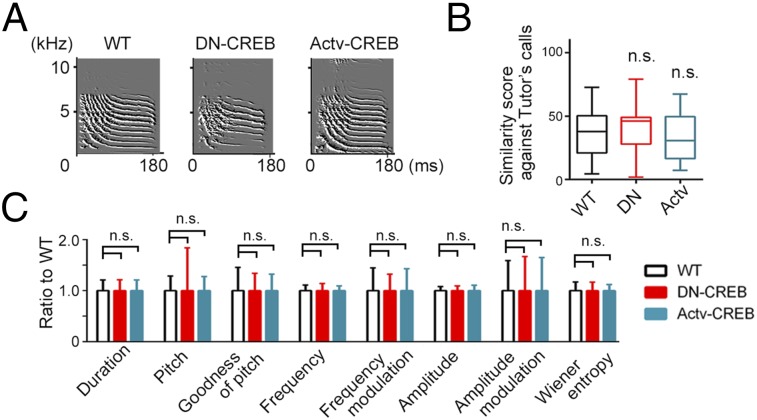
Songbirds postnatally develop their skill to utter and to perceive a vocal signal for communication. How genetic and environmental influences act in concert to regulate the development of such skill is not fully understood. Here, we report the phenotype of transgenic songbirds with altered intrinsic activity of cAMP response element-binding protein (CREB) transcription factor. By viral vector-mediated modification of genomic DNA, we established germ line-transmitted lines of zebra finches, which exhibited enhanced or suppressed activity of CREB. Although intrinsically acquired vocalizations or their hearing ability were not affected, the transgenic birds showed reduced vocal learning quality of their own songs and impaired audio-memory formation against conspecific songs. These results thus demonstrate that appropriate activity of CREB is necessary for the postnatal acquisition of learned behavior in songbirds, and the CREB transgenic birds offer a unique opportunity to separately manipulate both genetic and environmental factors that impinge on the postnatal song learning.
Keywords: CREB; postnatal development; songbird; transgenic animal; vocal learning.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Doupe AJ, Kuhl PK. Birdsong and human speech: Common themes and mechanisms. Annu Rev Neurosci. 1999;22:567–631. - PubMed
-
- Clayton DF. The genomics of memory and learning in songbirds. Annu Rev Genomics Hum Genet. 2013;14:45–65. - PubMed
-
- Konishi M. The role of auditory feedback in the control of vocalization in the white-crowned sparrow. Z Tierpsychol. 1965;22(7):770–783. - PubMed
-
- Marler P. A comparative approach to vocal learning: Song development in white-crowned sparrows. J Comp Physiol Psychol. 1970;71(2, Pt.2):1–25.
-
- Slater PJB. Bird song learning: Causes and consequences. Ethol Ecol Evol. 1989;1:19–46.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
