

Cdk1 Activates Pre-mitotic Nuclear Envelope Dynein Recruitment and Apical Nuclear Migration in Neural Stem Cells
- PMID: 26051540
- PMCID: PMC4480218
- DOI: 10.1016/j.devcel.2015.04.022
Cdk1 Activates Pre-mitotic Nuclear Envelope Dynein Recruitment and Apical Nuclear Migration in Neural Stem Cells
Abstract
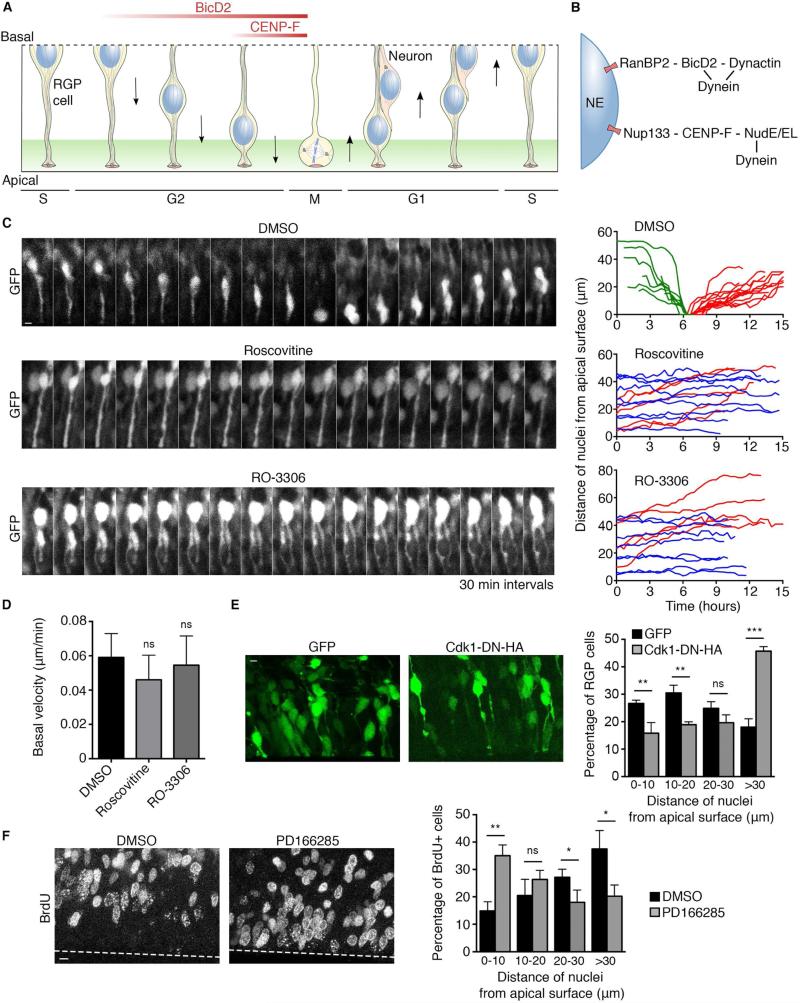
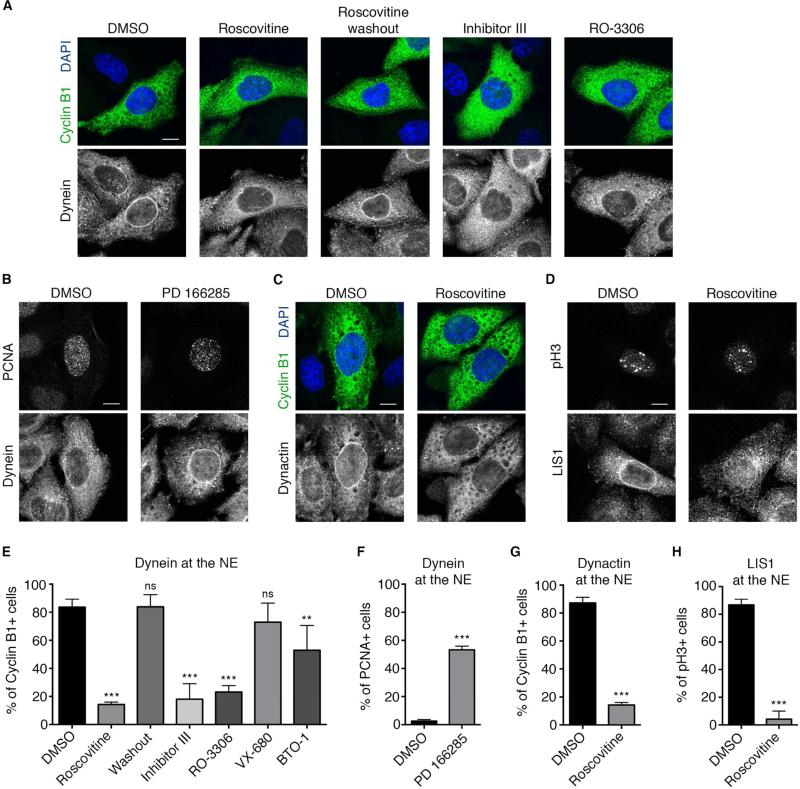
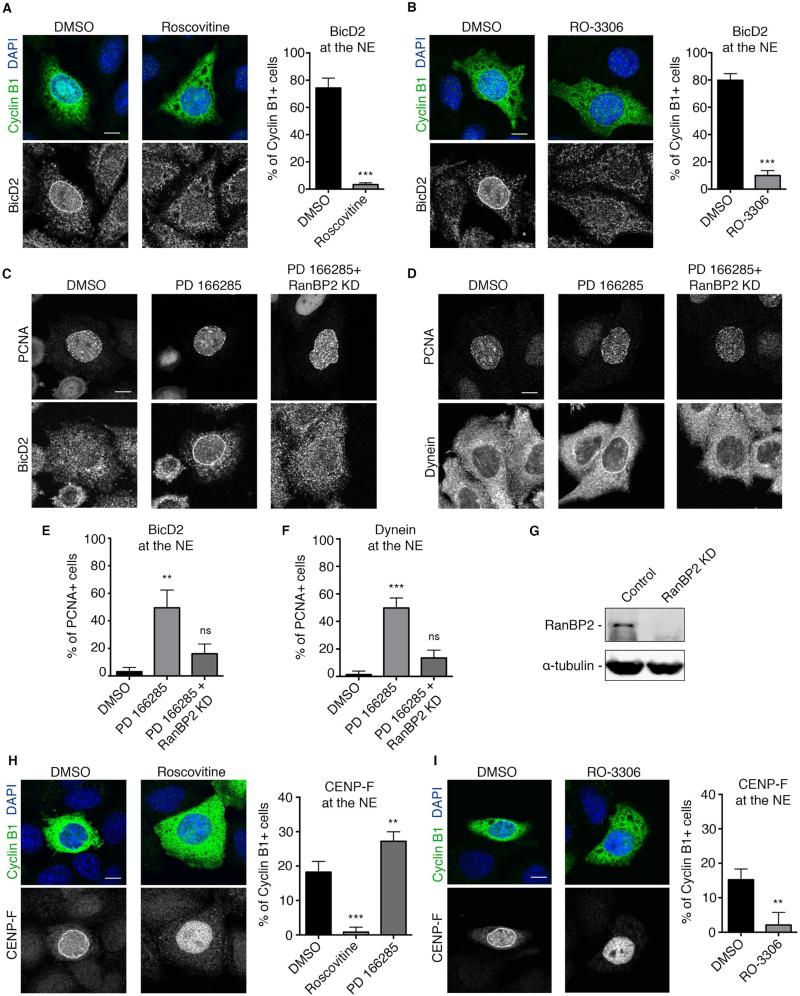
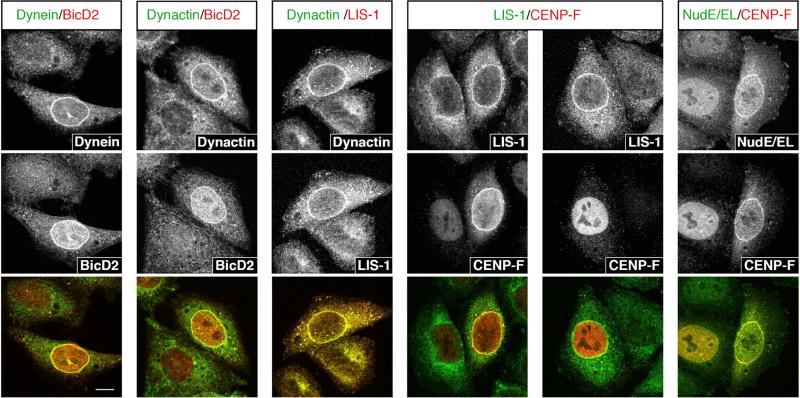
Dynein recruitment to the nuclear envelope is required for pre-mitotic nucleus-centrosome interactions in nonneuronal cells and for apical nuclear migration in neural stem cells. In each case, dynein is recruited to the nuclear envelope (NE) specifically during G2 via two nuclear pore-mediated mechanisms involving RanBP2-BicD2 and Nup133-CENP-F. The mechanisms responsible for cell-cycle control of this behavior are unknown. We now find that Cdk1 serves as a direct master controller for NE dynein recruitment in neural stem cells and HeLa cells. Cdk1 phosphorylates conserved sites within RanBP2 and activates BicD2 binding and early dynein recruitment. Late recruitment is triggered by a Cdk1-induced export of CENP-F from the nucleus. Forced NE targeting of BicD2 overrides Cdk1 inhibition, fully rescuing dynein recruitment and nuclear migration in neural stem cells. These results reveal how NE dynein recruitment is cell-cycle regulated and identify the trigger mechanism for apical nuclear migration in the brain.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
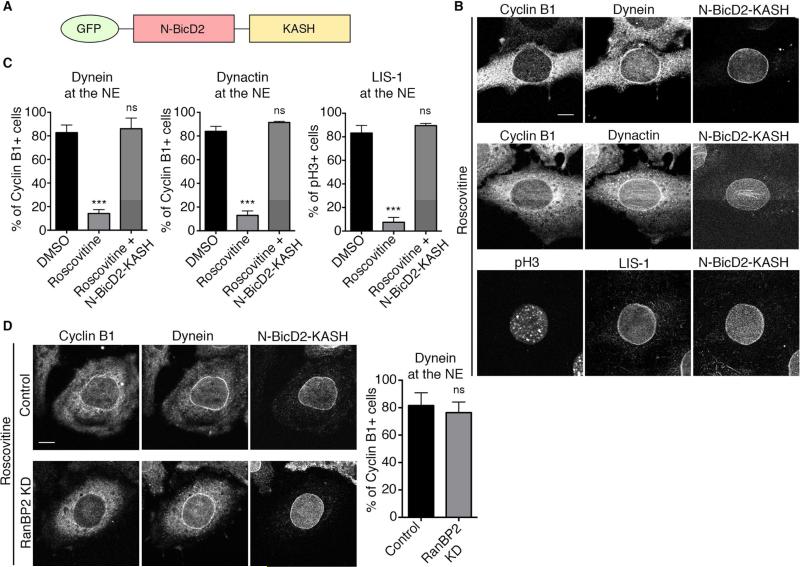
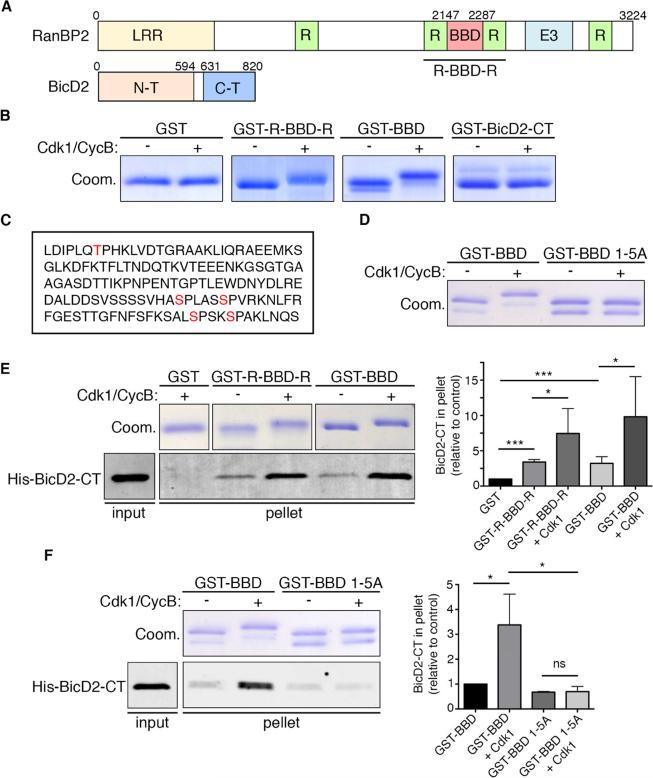
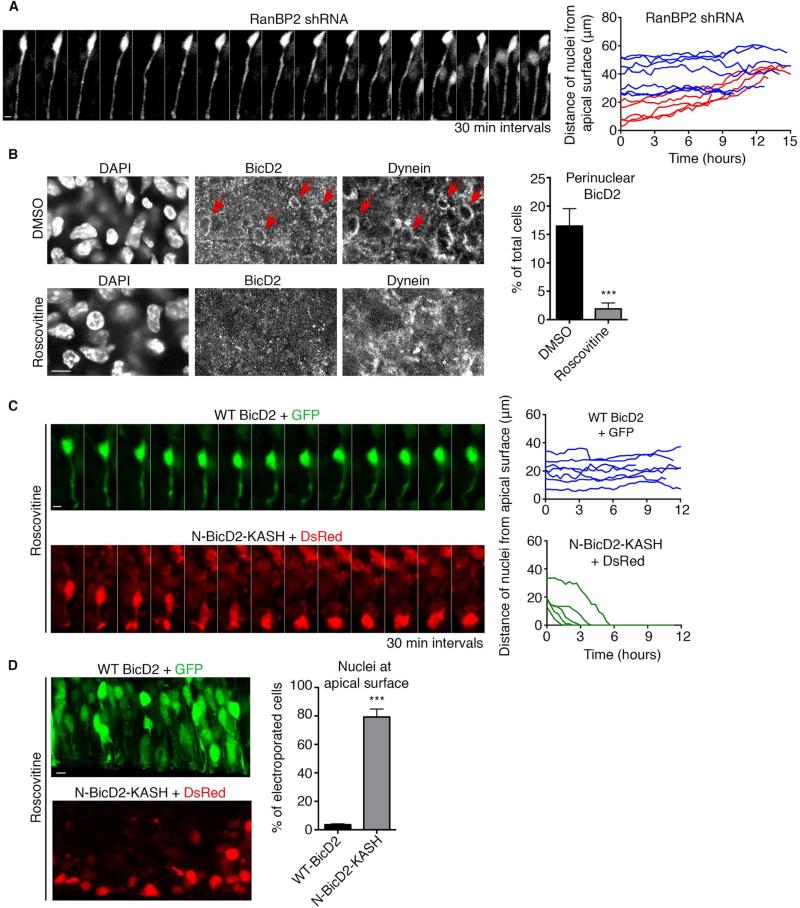
References
-
- Beaudouin J, Gerlich D, Daigle N, Eils R, Ellenberg J. Nuclear envelope breakdown proceeds by microtubule-induced tearing of the lamina. Cell. 2002;108:83–96. - PubMed
-
- Cai Y, Singh BB, Aslanukov A, Zhao H, Ferreira PA. The docking of kinesins, KIF5B and KIF5C, to Ran-binding protein 2 (RanBP2) is mediated via a novel RanBP2 domain. J Biol Chem. 2001;276:41594–41602. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
