

Quaternary Structure Defines a Large Class of Amyloid-β Oligomers Neutralized by Sequestration
- PMID: 26051935
- PMCID: PMC4494129
- DOI: 10.1016/j.celrep.2015.05.021
Quaternary Structure Defines a Large Class of Amyloid-β Oligomers Neutralized by Sequestration
Abstract
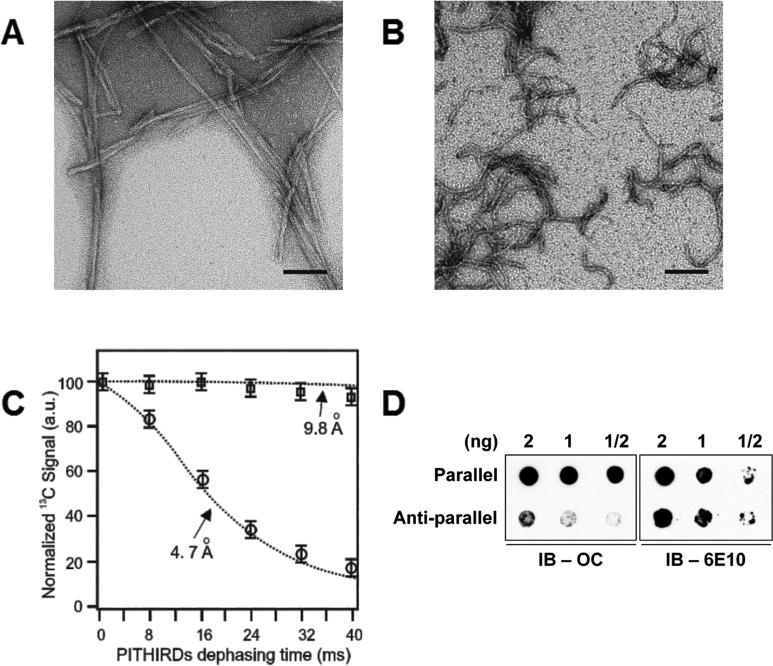
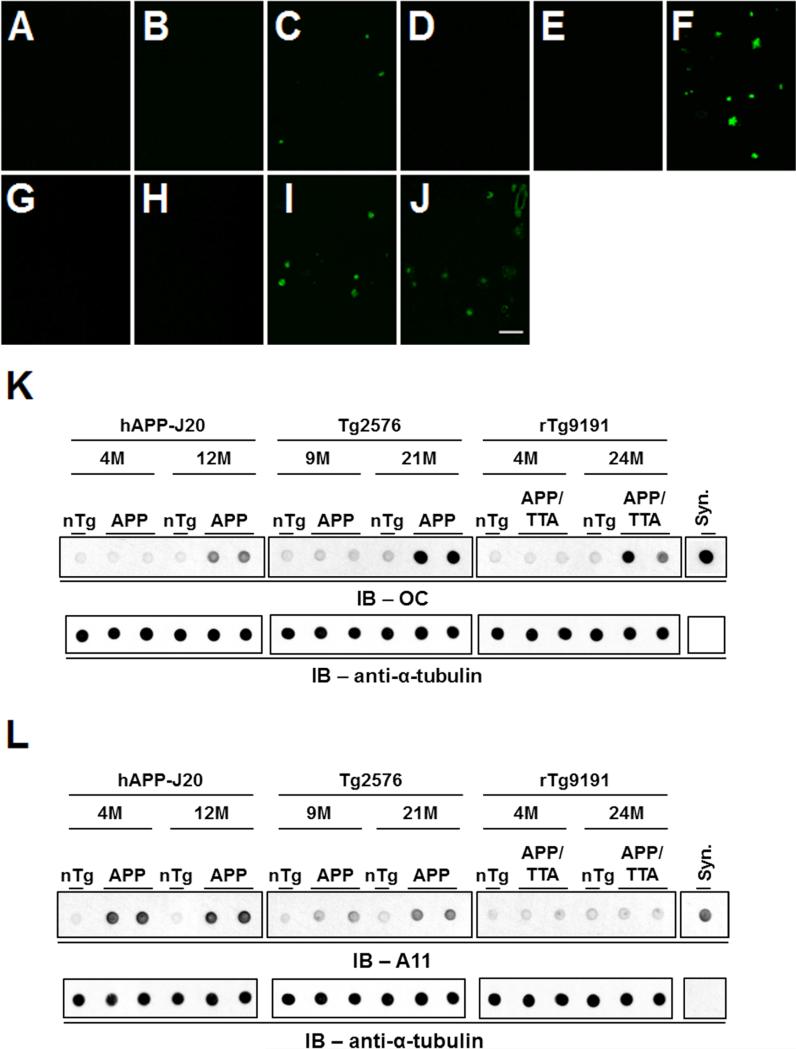
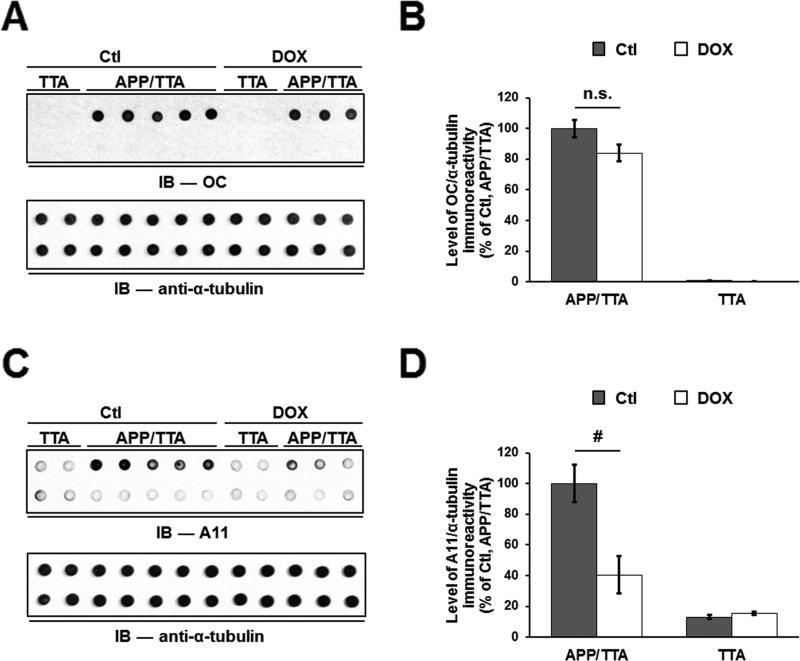
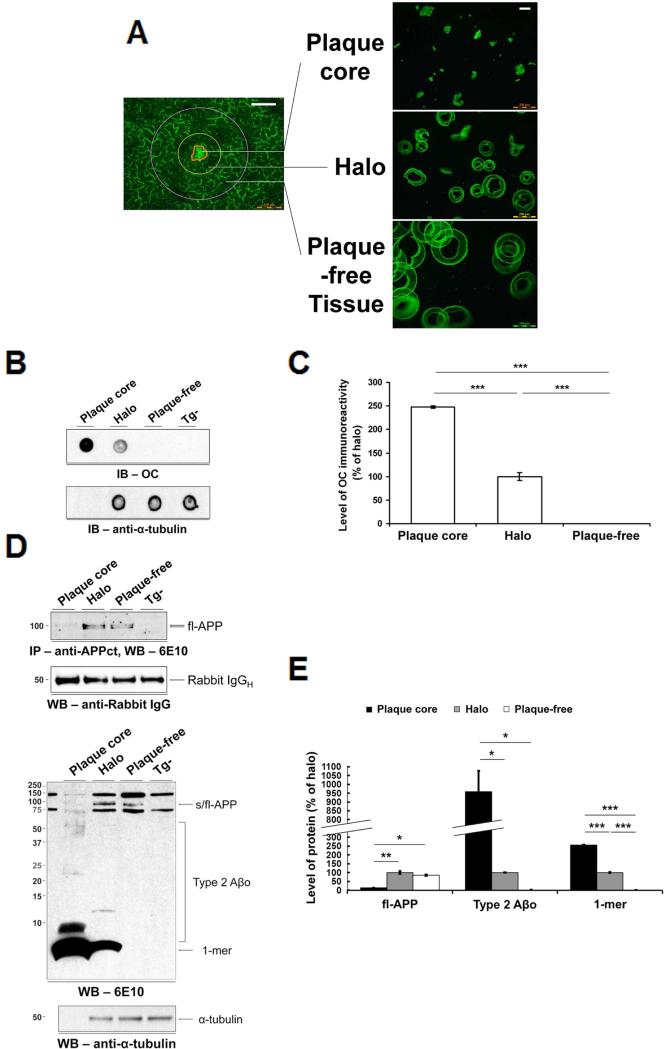
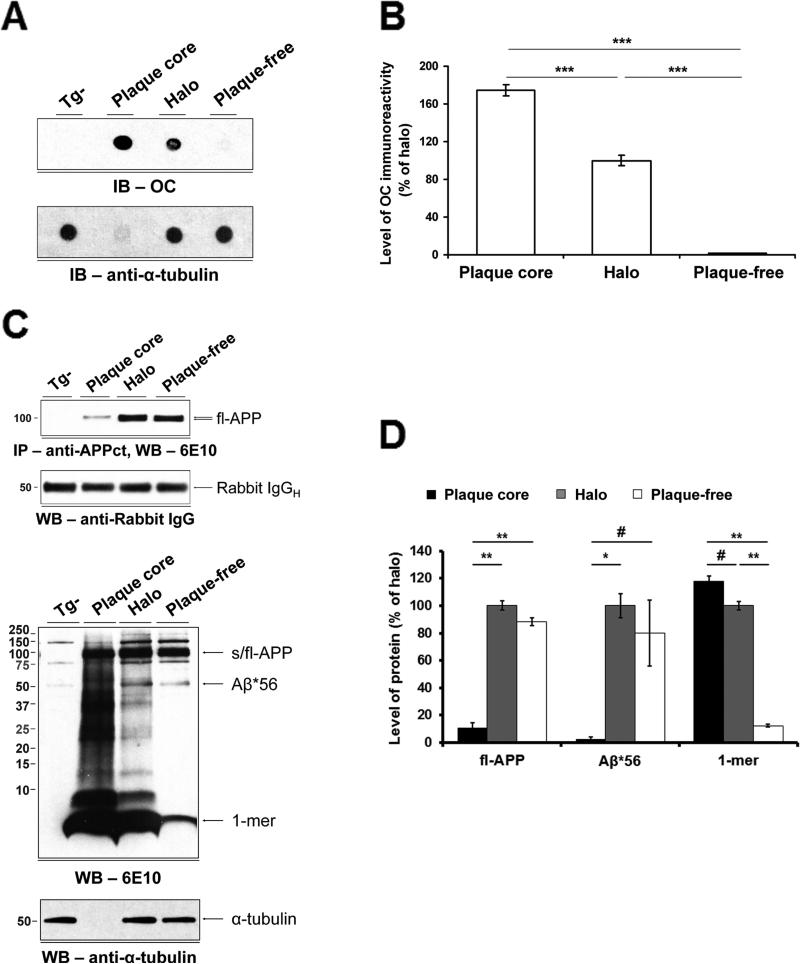
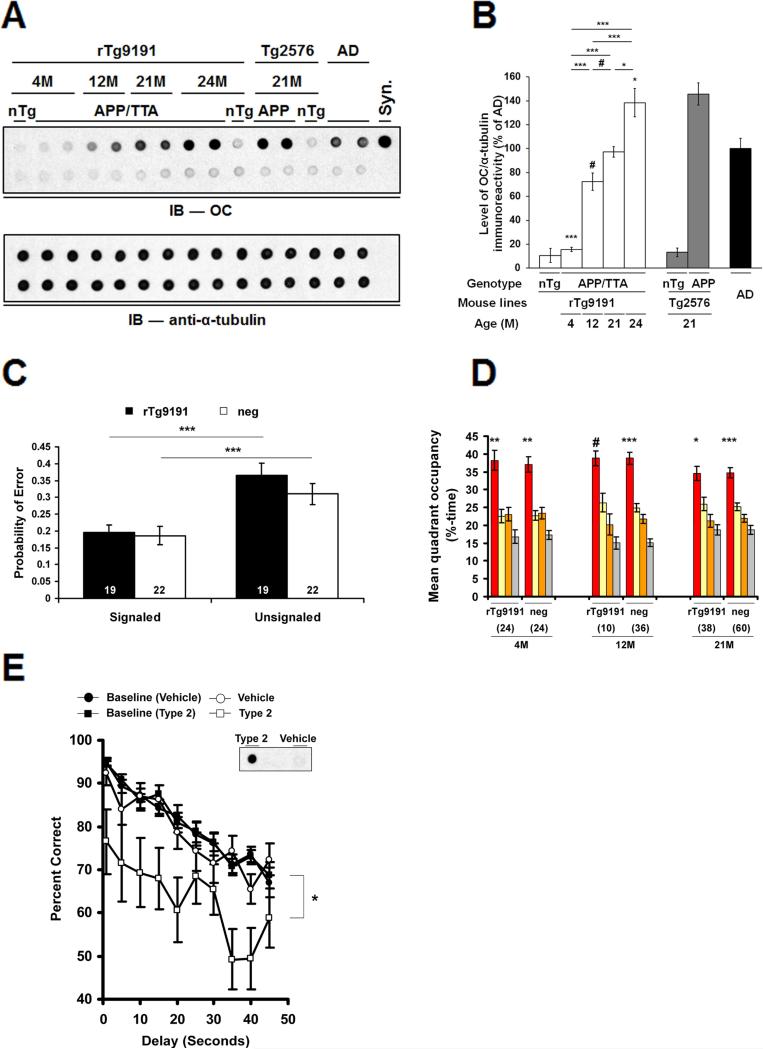
The accumulation of amyloid-β (Aβ) as amyloid fibrils and toxic oligomers is an important step in the development of Alzheimer's disease (AD). However, there are numerous potentially toxic oligomers and little is known about their neurological effects when generated in the living brain. Here we show that Aβ oligomers can be assigned to one of at least two classes (type 1 and type 2) based on their temporal, spatial, and structural relationships to amyloid fibrils. The type 2 oligomers are related to amyloid fibrils and represent the majority of oligomers generated in vivo, but they remain confined to the vicinity of amyloid plaques and do not impair cognition at levels relevant to AD. Type 1 oligomers are unrelated to amyloid fibrils and may have greater potential to cause global neural dysfunction in AD because they are dispersed. These results refine our understanding of the pathogenicity of Aβ oligomers in vivo.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Alzforum Bapineuzumab. 2015a
-
- Alzforum Gantenerumab. 2015b
-
- Benilova I, Karran E, De Strooper B. The toxic Abeta oligomer and Alzheimer's disease: an emperor in need of clothes. Nat Neurosci. 2012;15:349–357. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
