

Activity-Induced DNA Breaks Govern the Expression of Neuronal Early-Response Genes
- PMID: 26052046
- PMCID: PMC4886855
- DOI: 10.1016/j.cell.2015.05.032
Activity-Induced DNA Breaks Govern the Expression of Neuronal Early-Response Genes
Abstract
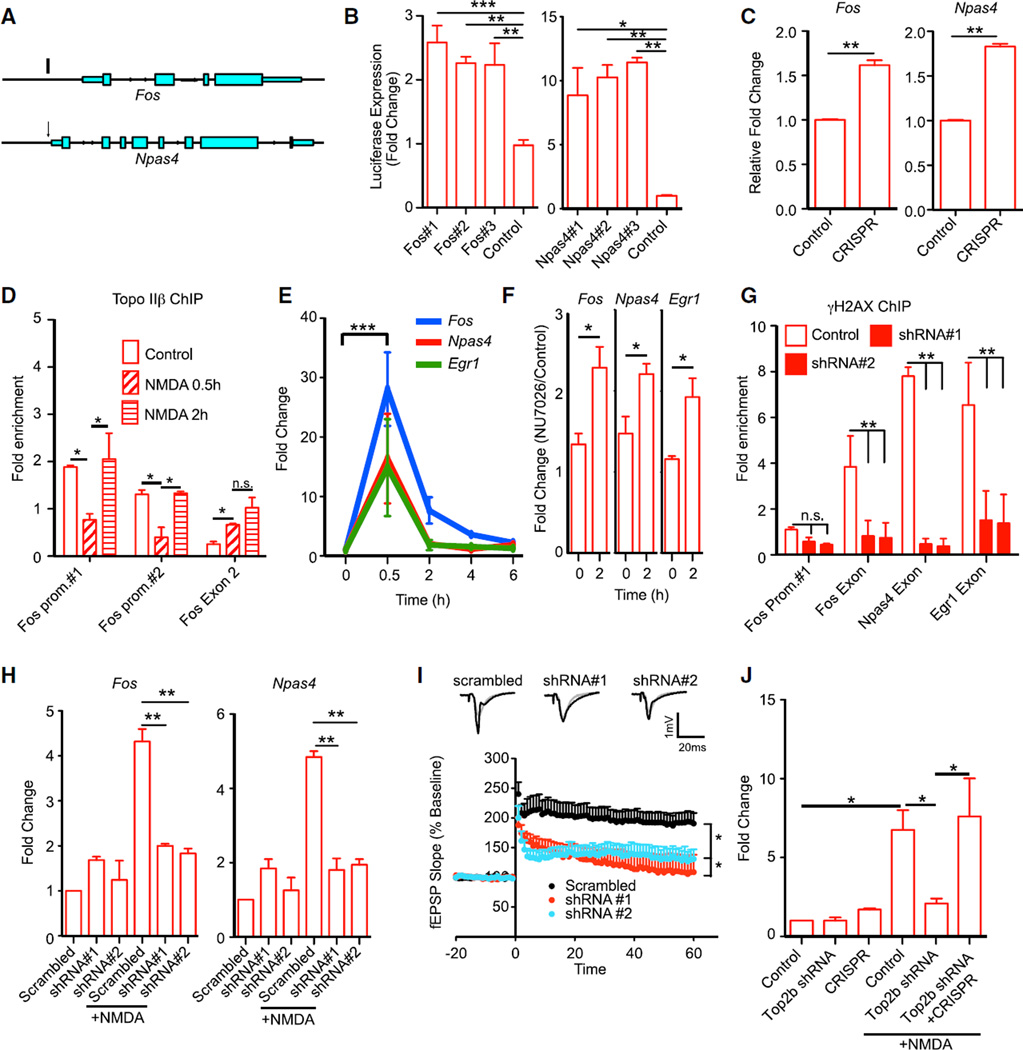
Neuronal activity causes the rapid expression of immediate early genes that are crucial for experience-driven changes to synapses, learning, and memory. Here, using both molecular and genome-wide next-generation sequencing methods, we report that neuronal activity stimulation triggers the formation of DNA double strand breaks (DSBs) in the promoters of a subset of early-response genes, including Fos, Npas4, and Egr1. Generation of targeted DNA DSBs within Fos and Npas4 promoters is sufficient to induce their expression even in the absence of an external stimulus. Activity-dependent DSB formation is likely mediated by the type II topoisomerase, Topoisomerase IIβ (Topo IIβ), and knockdown of Topo IIβ attenuates both DSB formation and early-response gene expression following neuronal stimulation. Our results suggest that DSB formation is a physiological event that rapidly resolves topological constraints to early-response gene expression in neurons.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures





Comment in
-
A Shortcut to Activity-Dependent Transcription.Cell. 2015 Jun 18;161(7):1496-8. doi: 10.1016/j.cell.2015.06.009. Cell. 2015. PMID: 26091031
-
DNA damage and repair regulate neuronal gene expression.Cell Res. 2015 Sep;25(9):993-4. doi: 10.1038/cr.2015.85. Epub 2015 Jul 17. Cell Res. 2015. PMID: 26183066 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
