

Schwann cell myelination
- PMID: 26054742
- PMCID: PMC4526746
- DOI: 10.1101/cshperspect.a020529
Schwann cell myelination
Abstract
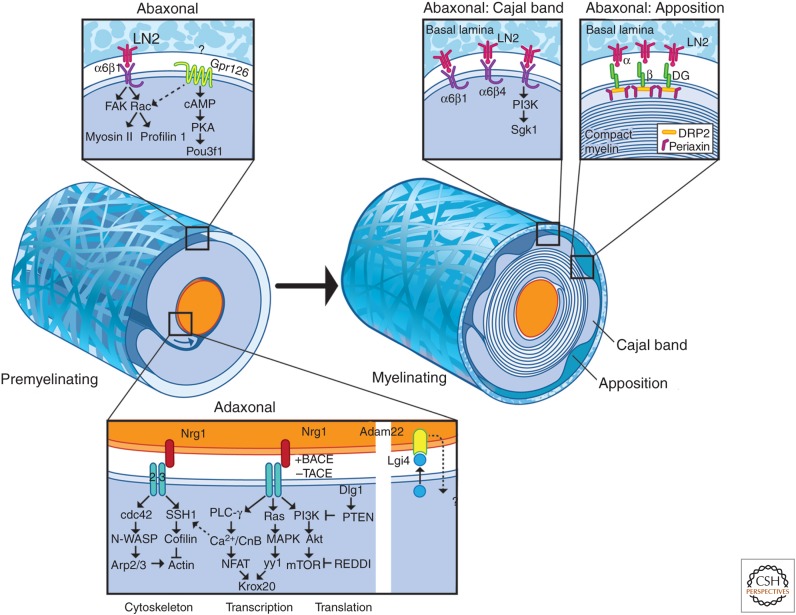
Myelinated nerve fibers are essential for the rapid propagation of action potentials by saltatory conduction. They form as the result of reciprocal interactions between axons and Schwann cells. Extrinsic signals from the axon, and the extracellular matrix, drive Schwann cells to adopt a myelinating fate, whereas myelination reorganizes the axon for its role in conduction and is essential for its integrity. Here, we review our current understanding of the development, molecular organization, and function of myelinating Schwann cells. Recent findings into the extrinsic signals that drive Schwann cell myelination, their cognate receptors, and the downstream intracellular signaling pathways they activate will be described. Together, these studies provide important new insights into how these pathways converge to activate the transcriptional cascade of myelination and remodel the actin cytoskeleton that is critical for morphogenesis of the myelin sheath.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
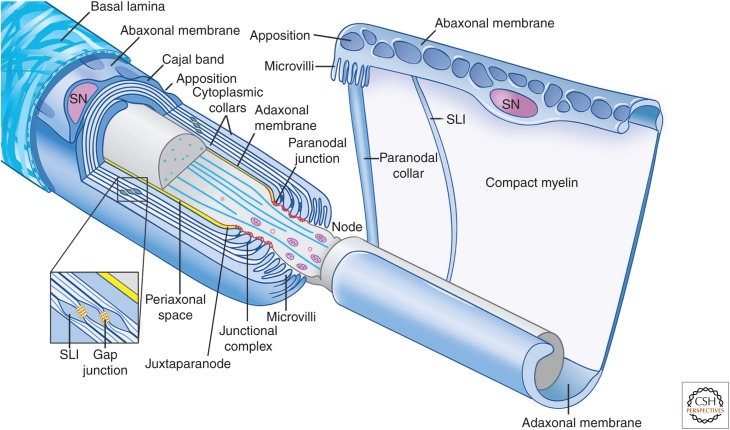
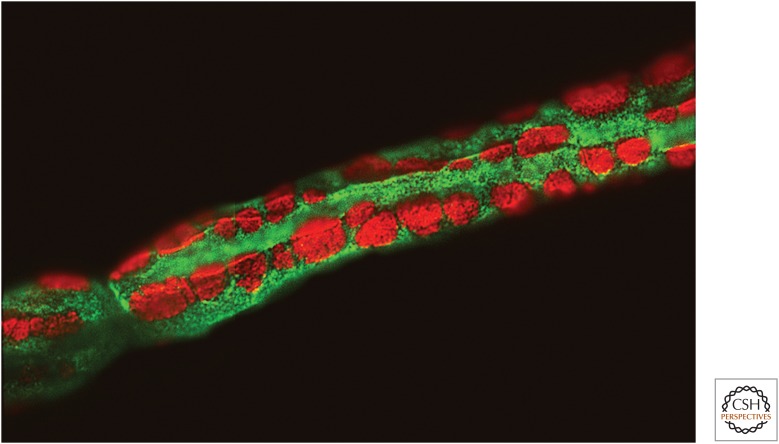
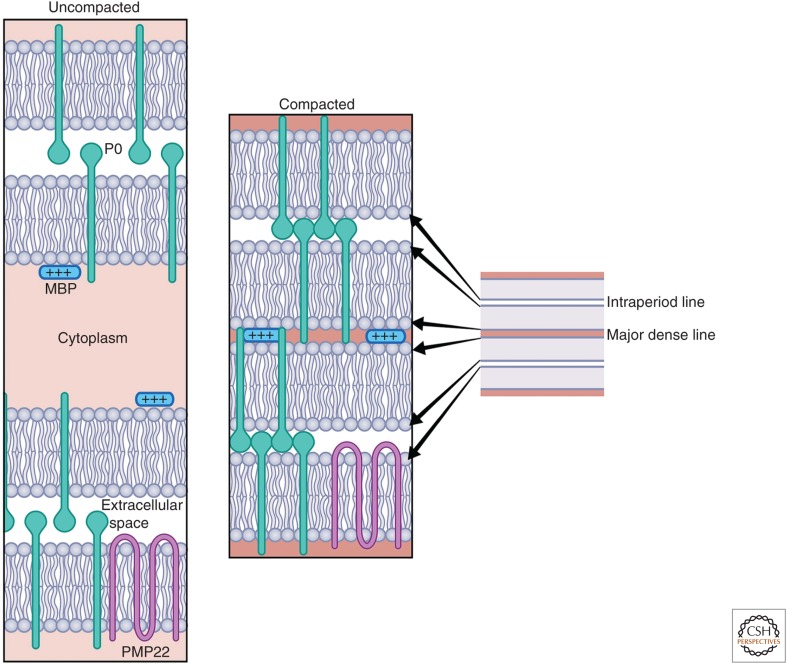
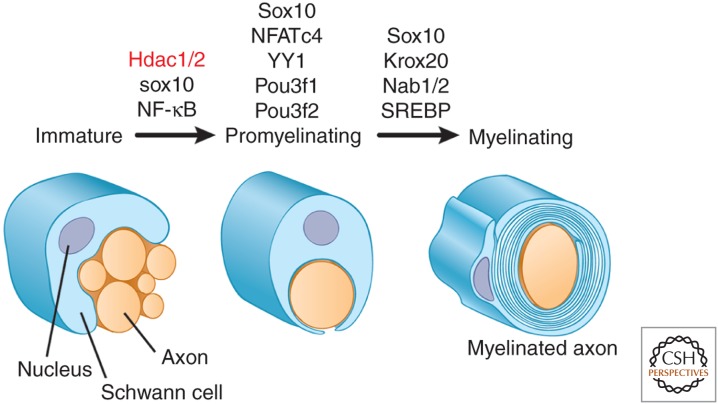
References
-
- Abe I, Ochiai N, Ichimura H, Tsujino A, Sun J, Hara Y. 2004. Internodes can nearly double in length with gradual elongation of the adult rat sciatic nerve. J Orthop Res 22: 571–577. - PubMed
-
- Aguayo AJ, Bray GM, Perkins SC. 1979. Axon-Schwann cell relationships in neuropathies of mutant mice. Ann NY Acad Sci 317: 512–531. - PubMed
-
- Armstrong R, Toews AD, Morell P. 1987. Axonal transport through nodes of Ranvier. Brain Res 412: 196–199. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources