

Smad7 Modulates Epidermal Growth Factor Receptor Turnover through Sequestration of c-Cbl
- PMID: 26055326
- PMCID: PMC4508311
- DOI: 10.1128/MCB.00274-15
Smad7 Modulates Epidermal Growth Factor Receptor Turnover through Sequestration of c-Cbl
Abstract
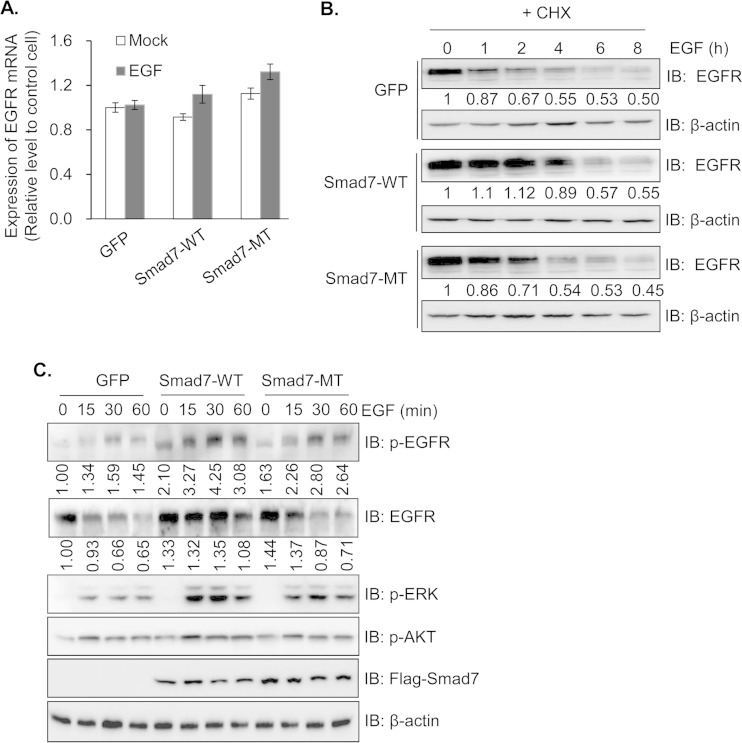
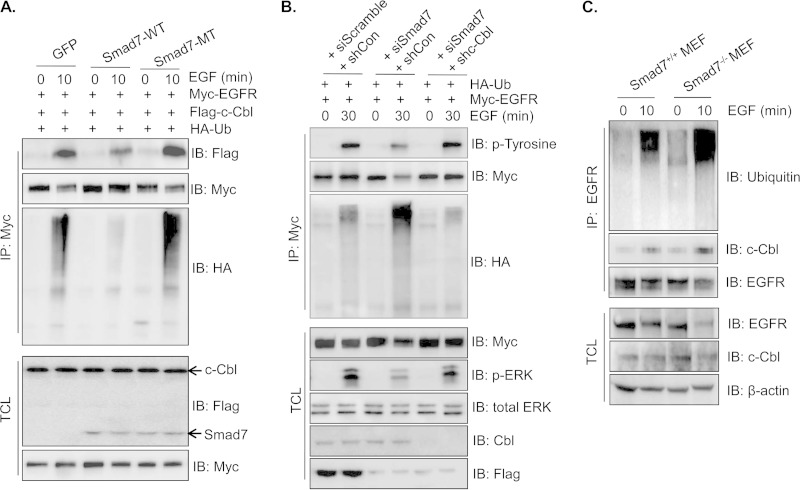
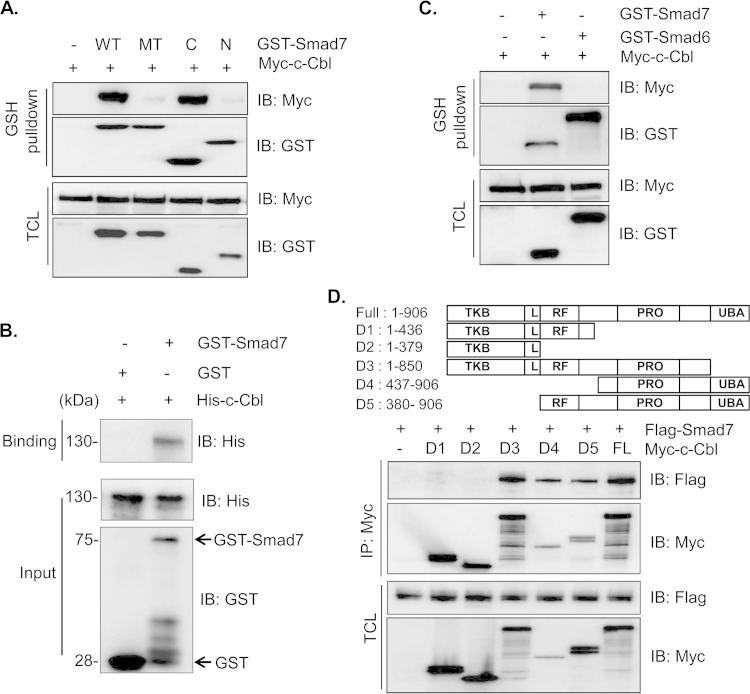
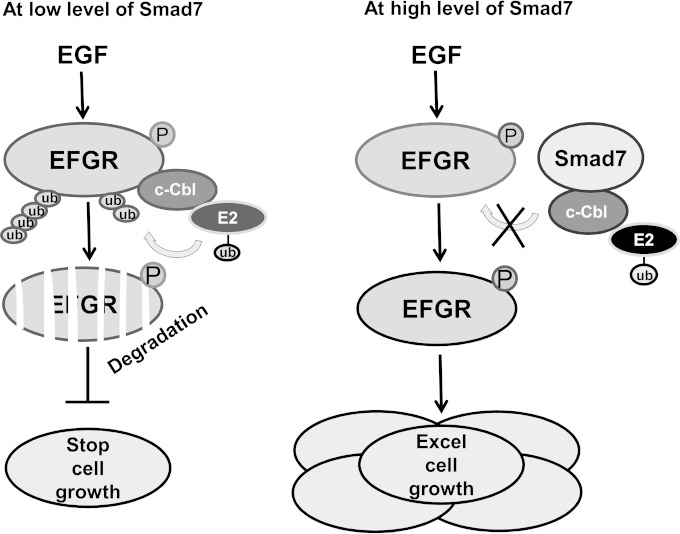
Epidermal growth factor (EGF) regulates various cellular events, including proliferation, differentiation, migration, and tumorigenesis. For the maintenance of homeostasis, EGF signaling should be tightly regulated to prevent the aberrant activation. Smad7 has been known as inhibitory Smad that blocks the signal transduction of transforming growth factor β. In the process of cell proliferation or transformation, Smad7 has been shown the opposite activities as a promoter or suppressor depending on cell types or microenvironments. We found that the overexpression of Smad7 in human HaCaT keratinocyte cells and mouse skin tissues elevated EGF receptor (EGFR) activity by impairing ligand-induced ubiquitination and degradation of activated receptor, which is induced by the E3 ubiquitin ligase c-Cbl. The C-terminal MH2 region but not MH1 region of Smad7 is critical for interaction with c-Cbl to inhibit the ubiquitination of EGFR. Interestingly, wild-type Smad7, but not Smad6 or mutant Smad7, destabilized the EGF-induced complex formation of c-Cbl and EGFR. These data suggest a novel role for Smad7 as a promoter for prolonging the EGFR signal in keratinocyte and skin tissue by reducing its ligand-induced ubiquitination and degradation.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous