

Insights into the evolution of enzyme substrate promiscuity after the discovery of (βα)₈ isomerase evolutionary intermediates from a diverse metagenome
- PMID: 26058375
- PMCID: PMC4462073
- DOI: 10.1186/s12862-015-0378-1
Insights into the evolution of enzyme substrate promiscuity after the discovery of (βα)₈ isomerase evolutionary intermediates from a diverse metagenome
Abstract
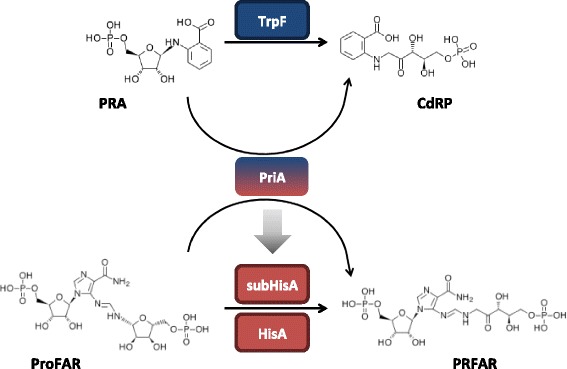
Background: Current sequence-based approaches to identify enzyme functional shifts, such as enzyme promiscuity, have proven to be highly dependent on a priori functional knowledge, hampering our ability to reconstruct evolutionary history behind these mechanisms. Hidden Markov Model (HMM) profiles, broadly used to classify enzyme families, can be useful to distinguish between closely related enzyme families with different specificities. The (βα)8-isomerase HisA/PriA enzyme family, involved in L-histidine (HisA, mono-substrate) biosynthesis in most bacteria and plants, but also in L-tryptophan (HisA/TrpF or PriA, dual-substrate) biosynthesis in most Actinobacteria, has been used as model system to explore evolutionary hypotheses and therefore has a considerable amount of evolutionary, functional and structural knowledge available. We searched for functional evolutionary intermediates between the HisA and PriA enzyme families in order to understand the functional divergence between these families.
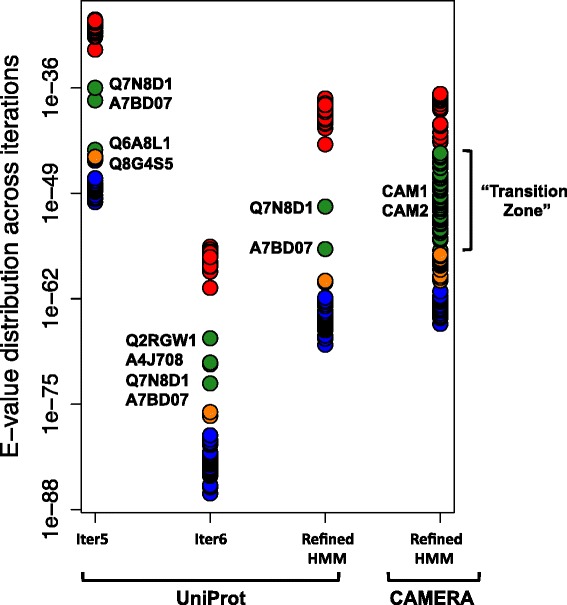
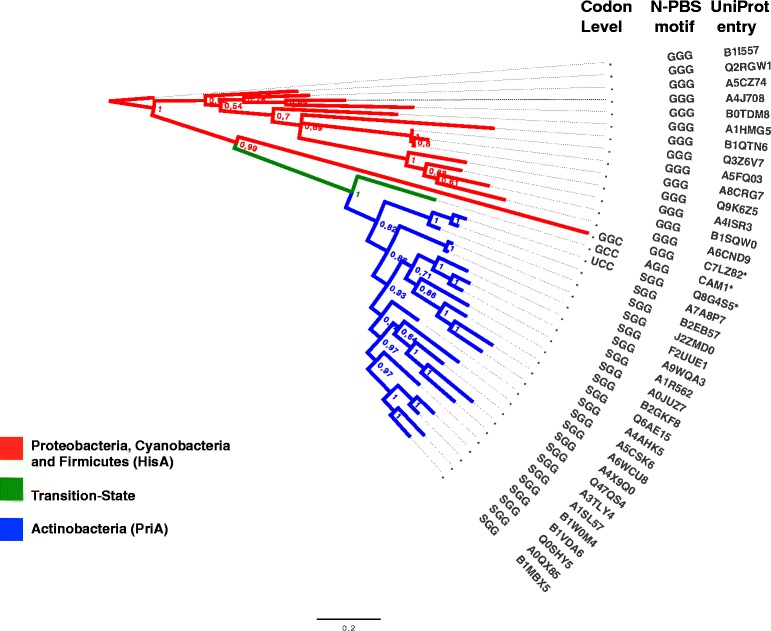
Results: We constructed a HMM profile that correctly classifies sequences of unknown function into the HisA and PriA enzyme sub-families. Using this HMM profile, we mined a large metagenome to identify plausible evolutionary intermediate sequences between HisA and PriA. These sequences were used to perform phylogenetic reconstructions and to identify functionally conserved amino acids. Biochemical characterization of one selected enzyme (CAM1) with a mutation within the functionally essential N-terminus phosphate-binding site, namely, an alanine instead of a glycine in HisA or a serine in PriA, showed that this evolutionary intermediate has dual-substrate specificity. Moreover, site-directed mutagenesis of this alanine residue, either backwards into a glycine or forward into a serine, revealed the robustness of this enzyme. None of these mutations, presumably upon functionally essential amino acids, significantly abolished its enzyme activities. A truncated version of this enzyme (CAM2) predicted to adopt a (βα)6-fold, and thus entirely lacking a C-terminus phosphate-binding site, was identified and shown to have HisA activity.
Conclusion: As expected, reconstruction of the evolution of PriA from HisA with HMM profiles suggest that functional shifts involve mutations in evolutionarily intermediate enzymes of otherwise functionally essential residues or motifs. These results are in agreement with a link between promiscuous enzymes and intragenic epistasis. HMM provides a convenient approach for gaining insights into these evolutionary processes.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
