

Critical role of the α1-Na(+), K(+)-ATPase subunit in insensitivity of rodent cells to cytotoxic action of ouabain
- PMID: 26067145
- PMCID: PMC4677477
- DOI: 10.1007/s10495-015-1144-y
Critical role of the α1-Na(+), K(+)-ATPase subunit in insensitivity of rodent cells to cytotoxic action of ouabain
Abstract
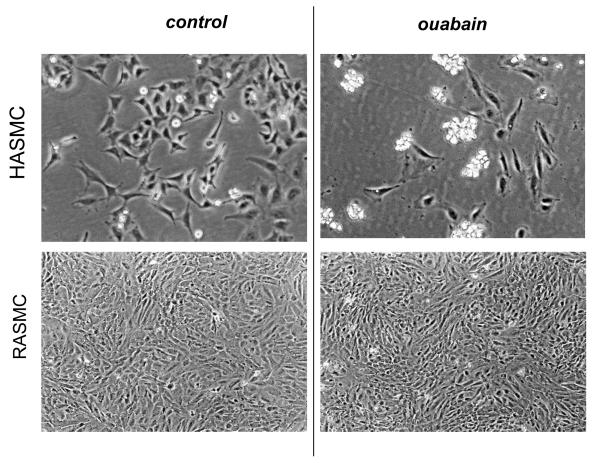
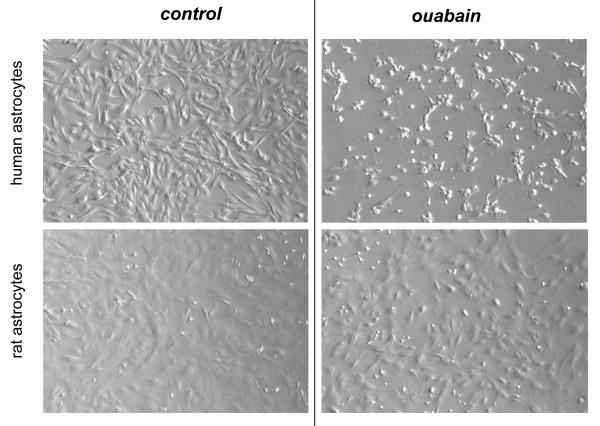
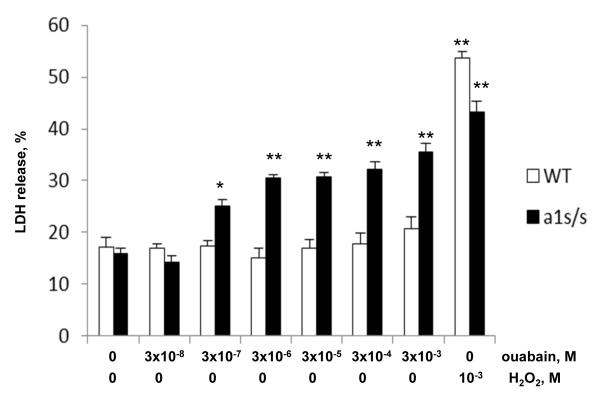
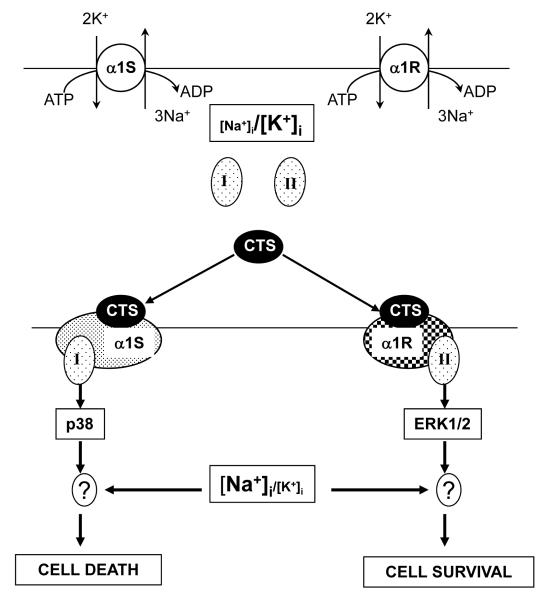
In rodents, ubiquitous α1-Na(+), K(+)-ATPase is inhibited by ouabain and other cardiotonic steroids (CTS) at ~10(3)-fold higher concentrations than those effective in other mammals. To examine the specific roles of the CTS-sensitive α1S- and CTS-resistant α1R-Na(+), K(+)-ATPase isoforms, we compared the effects of ouabain on intracellular Na(+) and K(+) content, cell survival, and mitogen-activated protein kinases (MAPK) in human and rat vascular smooth muscle cells (HASMC and RASMC), human and rat endothelial cells (HUVEC and RAEC), and human and rat brain astrocytes. 6-h exposure of HASMC and HUVEC to 3 μM ouabain dramatically increased the intracellular [Na(+)]/[K(+)] ratio to the same extend as in RASMC and RAEC treated with 3000 μM ouabain. In 24, 3 μM ouabain triggered the death of all types of human cells used in this study. Unlike human cells, we did not detect any effect of 3000-5000 μM ouabain on the survival of rat cells, or smooth muscle cells from mouse aorta (MASMC). Unlike in the wild-type α1(R/R) mouse, ouabain triggered death of MASMC from α1(S/S) mouse expressing human α1-Na(+), K(+)-ATPase. Furthermore, transfection of HUVEC with rat α1R-Na(+), K(+)-ATPase protected them from the ouabain-induced death. In HUVEC, ouabain led to phosphorylation of p38 MAPK, whereas in RAEC it stimulated phosphorylation of ERK1/2. Overall, our results, demonstrate that the drastic differences in cytotoxic action of ouabain on human and rodent cells are caused by unique features of α1S/α1R-Na(+), K(+)-ATPase, rather than by any downstream CTS-sensitive/resistant components of the cell death machinery.
Figures



References
-
- Therien AG, Blostien R. Mechanisms of sodium pump regulation. Am.J.Physiol. 2000;279:C541–C566. - PubMed
-
- Scheiner-Bobis G. The sodium pump. Its molecular properties and mechanisc of ion transport. Eur.J.Biochem. 2002;269:2424–2433. - PubMed
-
- Schoner W, Scheiner-Bobis G. Endogenous and exogenous cardiac glycosides: their role in hypertension, salt metabolism, and cell growth. Am.J.Physiol.Cell Physiol. 2007;293:C509–C536. - PubMed
-
- Lingrel JB, Argьello JM, Van Huysse JW, Kuntzweiler TA. Cation and cardiac glycoside binding sites of the Na,K-ATPase. Ann.N.Y.Acad.Sci. 1997;843:194–206. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
